

Distinct Transcriptional Programs Control Cross-Priming in Classical and Monocyte-Derived Dendritic Cells
- PMID: 27264183
- PMCID: PMC4941620
- DOI: 10.1016/j.celrep.2016.05.025
Distinct Transcriptional Programs Control Cross-Priming in Classical and Monocyte-Derived Dendritic Cells
Abstract
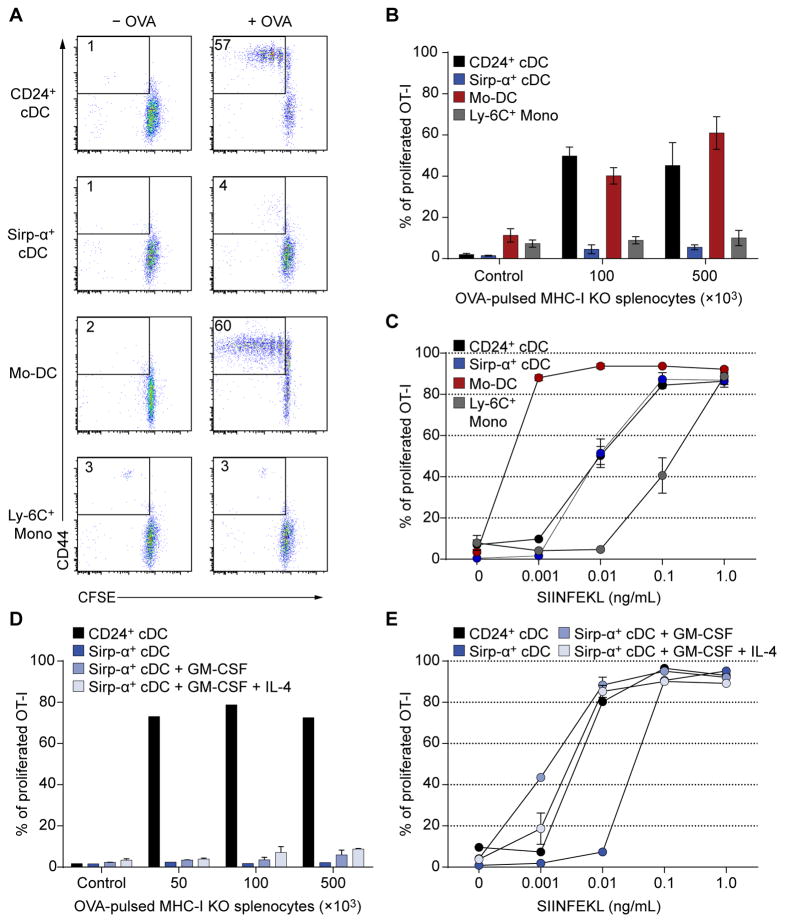
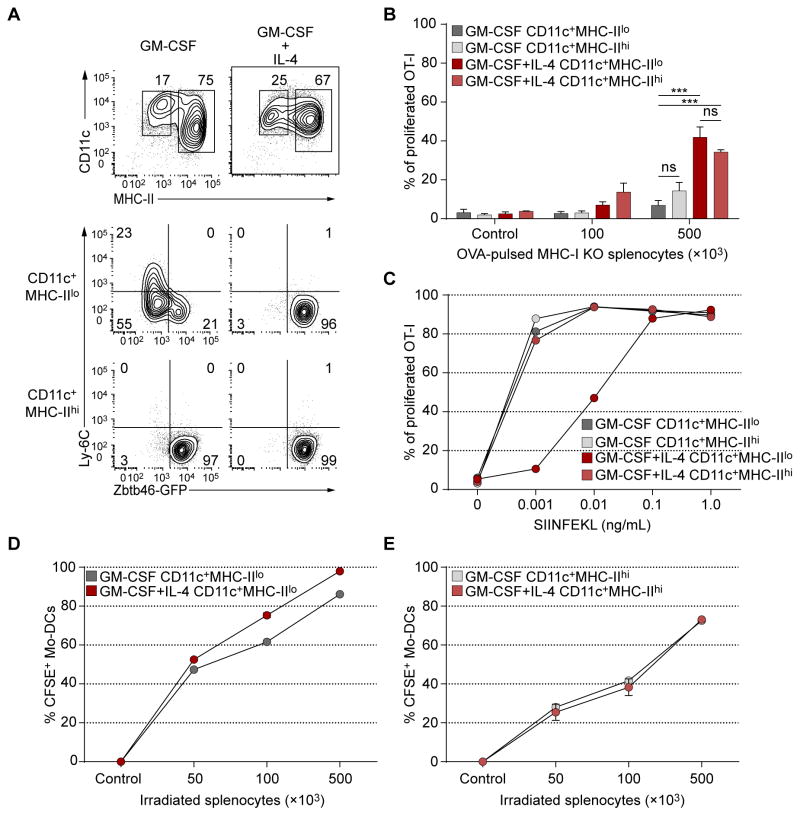
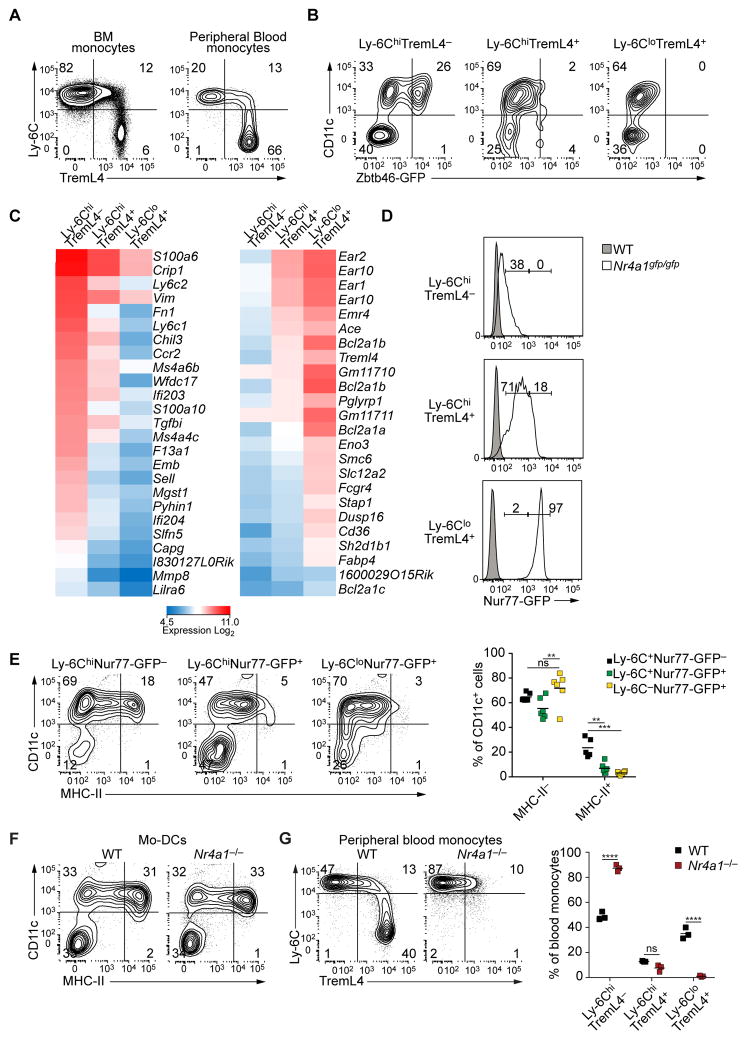
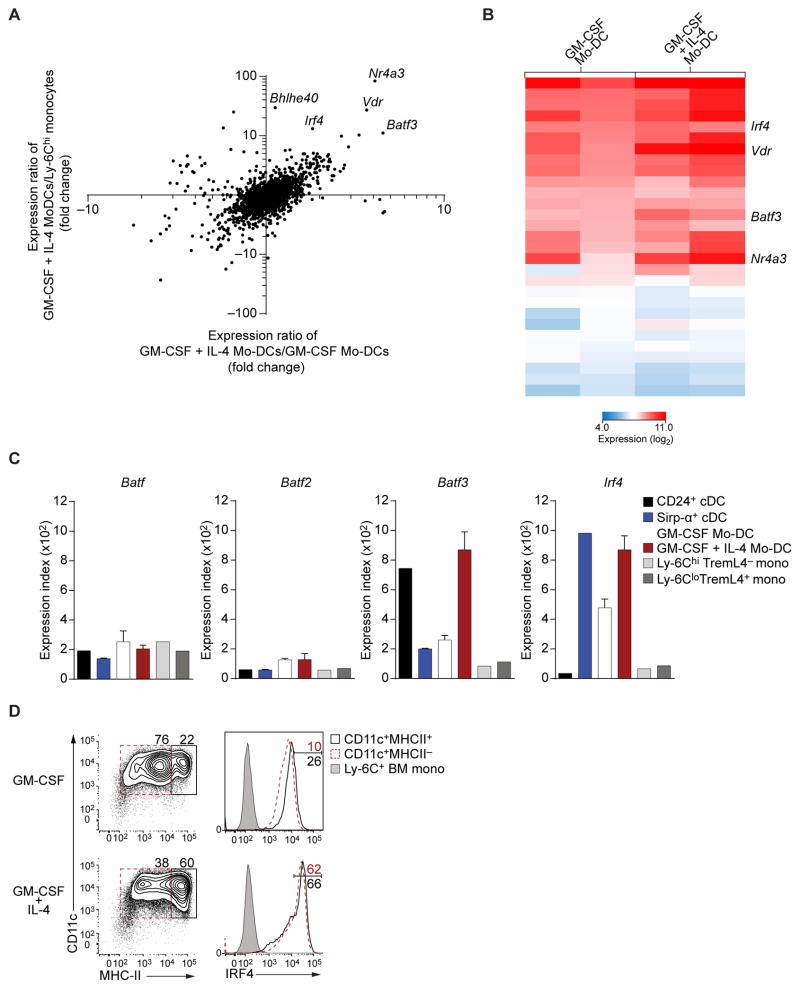
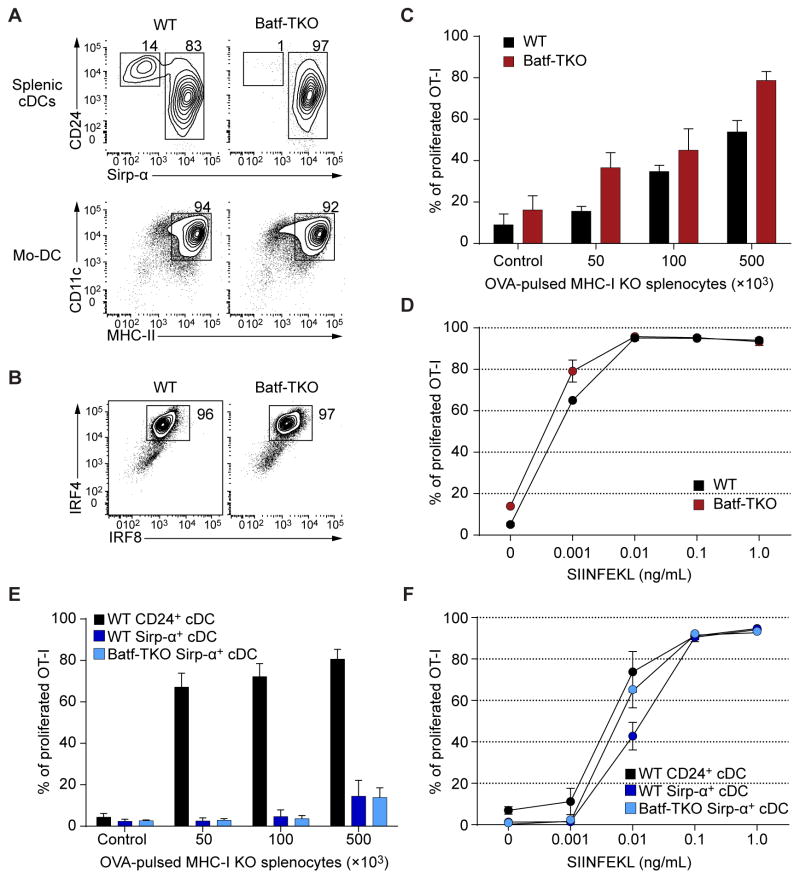
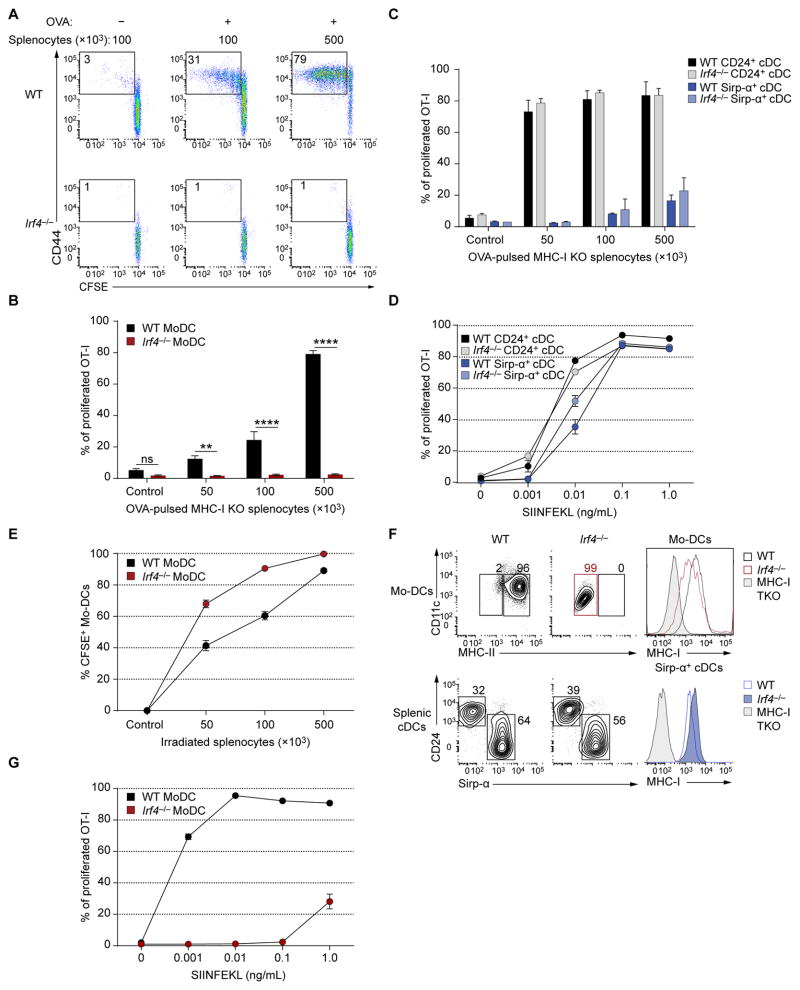
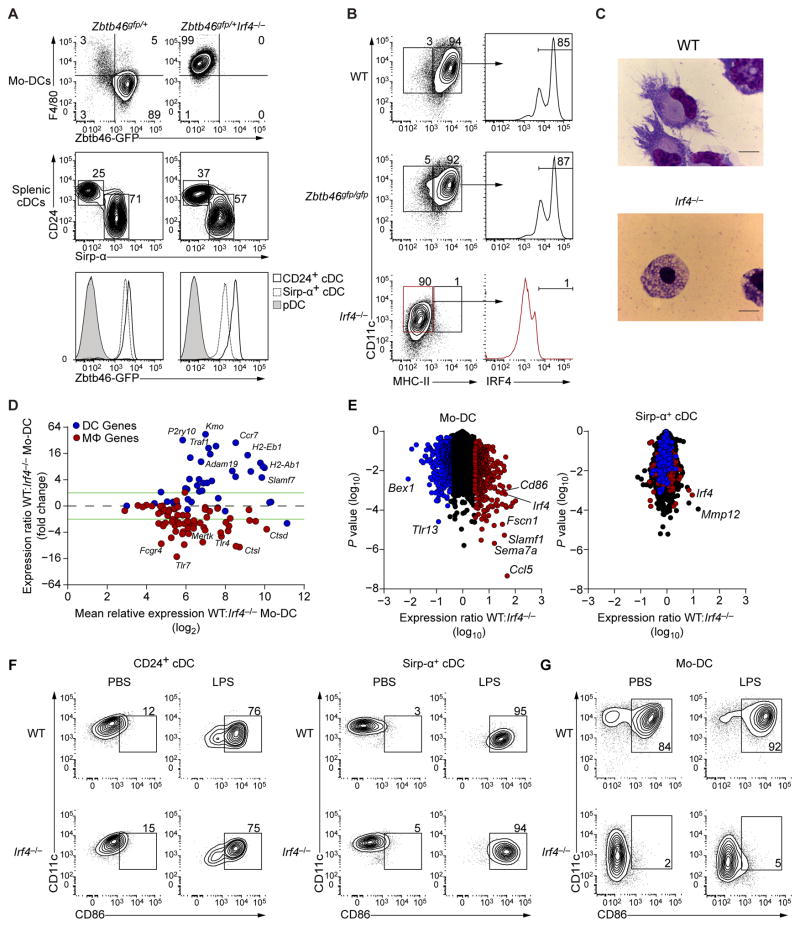
Both classical DCs (cDCs) and monocyte-derived DCs (Mo-DCs) are capable of cross-priming CD8(+) T cells in response to cell-associated antigens. We found that Ly-6C(hi)TREML4(-) monocytes can differentiate into Zbtb46(+) Mo-DCs in response to granulocyte-macrophage colony-stimulating factor (GM-CSF) and interleukin-4 (IL-4) but that Ly-6C(hi)TREML4(+) monocytes were committed to differentiate into Ly-6C(lo)TREML4(+) monocytes. Differentiation of Zbtb46(+) Mo-DCs capable of efficient cross-priming required both GM-CSF and IL-4 and was accompanied by the induction of Batf3 and Irf4. However, monocytes require IRF4, but not BATF3, to differentiate into Zbtb46(+) Mo-DCs capable of cross-priming CD8(+) T cells. Instead, Irf4(-/-) monocytes differentiate into macrophages in response to GM-CSF and IL-4. Thus, cDCs and Mo-DCs require distinct transcriptional programs of differentiation in acquiring the capacity to prime CD8(+) T cells. These differences may be of consideration in the use of therapeutic DC vaccines based on Mo-DCs.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Becker M, Guttler S, Bachem A, Hartung E, Mora A, Jakel A, Hutloff A, Henn V, Mages HW, Gurka S, Kroczek RA. Ontogenic, Phenotypic, and Functional Characterization of XCR1(+) Dendritic Cells Leads to a Consistent Classification of Intestinal Dendritic Cells Based on the Expression of XCR1 and SIRPalpha. Front Immunol. 2014;5:326. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
