

Morphomechanical Innovation Drives Explosive Seed Dispersal
- PMID: 27264605
- PMCID: PMC4930488
- DOI: 10.1016/j.cell.2016.05.002
Morphomechanical Innovation Drives Explosive Seed Dispersal
Abstract
How mechanical and biological processes are coordinated across cells, tissues, and organs to produce complex traits is a key question in biology. Cardamine hirsuta, a relative of Arabidopsis thaliana, uses an explosive mechanism to disperse its seeds. We show that this trait evolved through morphomechanical innovations at different spatial scales. At the organ scale, tension within the fruit wall generates the elastic energy required for explosion. This tension is produced by differential contraction of fruit wall tissues through an active mechanism involving turgor pressure, cell geometry, and wall properties of the epidermis. Explosive release of this tension is controlled at the cellular scale by asymmetric lignin deposition within endocarp b cells-a striking pattern that is strictly associated with explosive pod shatter across the Brassicaceae plant family. By bridging these different scales, we present an integrated mechanism for explosive seed dispersal that links evolutionary novelty with complex trait innovation. VIDEO ABSTRACT.
Copyright © 2016 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures


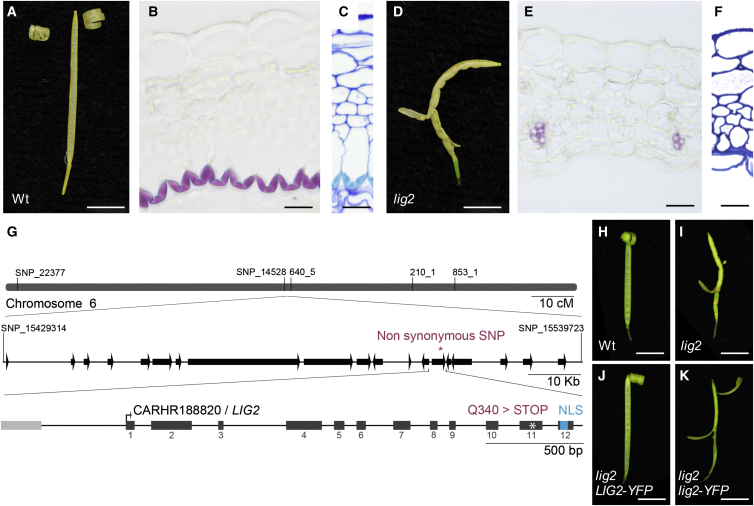


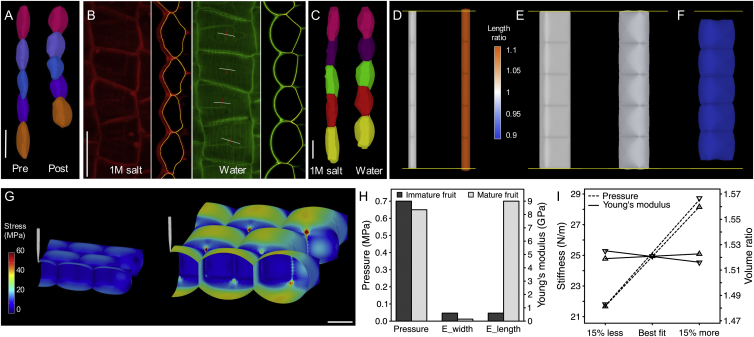

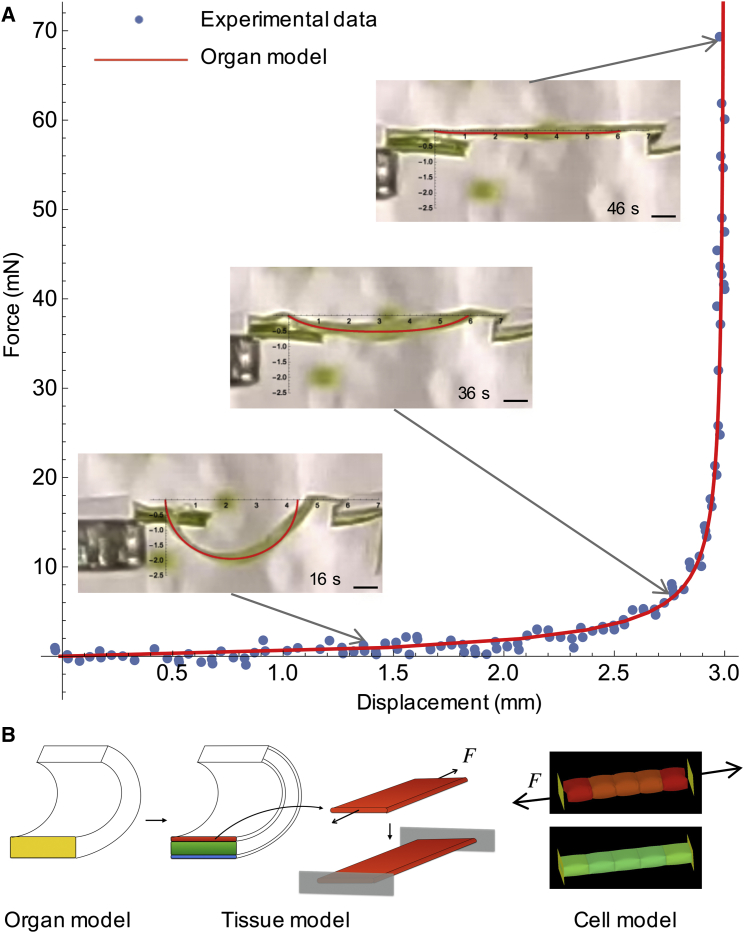
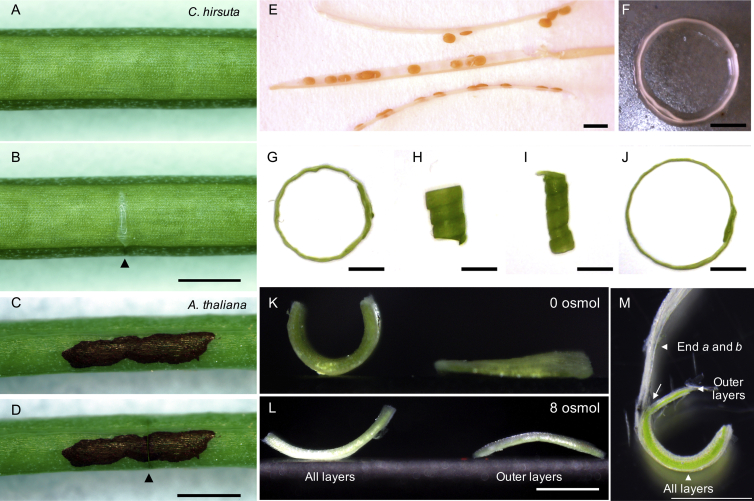






Comment in
-
Actuators Acting without Actin.Cell. 2016 Jun 30;166(1):15-7. doi: 10.1016/j.cell.2016.06.030. Cell. 2016. PMID: 27368097
Similar articles
-
Explosive seed dispersal depends on SPL7 to ensure sufficient copper for localized lignin deposition via laccases.Proc Natl Acad Sci U S A. 2022 Jun 14;119(24):e2202287119. doi: 10.1073/pnas.2202287119. Epub 2022 Jun 6. Proc Natl Acad Sci U S A. 2022. PMID: 35666865 Free PMC article.
-
Creating an explosion: Form and function in explosive fruit.Curr Opin Plant Biol. 2024 Jun;79:102543. doi: 10.1016/j.pbi.2024.102543. Epub 2024 Apr 30. Curr Opin Plant Biol. 2024. PMID: 38688200 Review.
-
The mechanism for explosive seed dispersal in Cardamine hirsuta (Brassicaceae).Am J Bot. 2011 Aug;98(8):1276-85. doi: 10.3732/ajb.1000374. Epub 2011 Jul 27. Am J Bot. 2011. PMID: 21795731
-
Explosive seed dispersal.New Phytol. 2017 Oct;216(2):339-342. doi: 10.1111/nph.14541. Epub 2017 Mar 20. New Phytol. 2017. PMID: 28318011 No abstract available.
-
Drawing lines and borders: how the dehiscent fruit of Arabidopsis is patterned.Bioessays. 2005 Jan;27(1):42-9. doi: 10.1002/bies.20165. Bioessays. 2005. PMID: 15612035 Review.
Cited by
-
Explosive seed dispersal depends on SPL7 to ensure sufficient copper for localized lignin deposition via laccases.Proc Natl Acad Sci U S A. 2022 Jun 14;119(24):e2202287119. doi: 10.1073/pnas.2202287119. Epub 2022 Jun 6. Proc Natl Acad Sci U S A. 2022. PMID: 35666865 Free PMC article.
-
Plant science's next top models.Ann Bot. 2020 Jun 19;126(1):1-23. doi: 10.1093/aob/mcaa063. Ann Bot. 2020. PMID: 32271862 Free PMC article. Review.
-
A DOF transcriptional repressor-gibberellin feedback loop plays a crucial role in modulating light-independent seed germination.Plant Commun. 2025 Apr 14;6(4):101262. doi: 10.1016/j.xplc.2025.101262. Epub 2025 Jan 28. Plant Commun. 2025. PMID: 39876558 Free PMC article.
-
Convergent Evolution of the Seed Shattering Trait.Genes (Basel). 2019 Jan 19;10(1):68. doi: 10.3390/genes10010068. Genes (Basel). 2019. PMID: 30669440 Free PMC article. Review.
-
Dehydration-induced corrugated folding in Rhapis excelsa plant leaves.Proc Natl Acad Sci U S A. 2024 Apr 23;121(17):e2320259121. doi: 10.1073/pnas.2320259121. Epub 2024 Apr 8. Proc Natl Acad Sci U S A. 2024. PMID: 38588439 Free PMC article.
References
-
- Barkoulas M., Hay A., Kougioumoutzi E., Tsiantis M. A developmental framework for dissected leaf formation in the Arabidopsis relative Cardamine hirsuta. Nat. Genet. 2008;40:1136–1141. - PubMed
-
- Bassel G.W., Stamm P., Mosca G., Barbier de Reuille P., Gibbs D.J., Winter R., Janka A., Holdsworth M.J., Smith R.S. Mechanical constraints imposed by 3D cellular geometry and arrangement modulate growth patterns in the Arabidopsis embryo. Proc. Natl. Acad. Sci. USA. 2014;111:8685–8690. - PMC - PubMed
-
- Beer T., Swaine M.D. On the theory of explosively dispersed seeds. New Phytol. 1977;78:681–694. - PubMed
