

Identification of ETV6-RUNX1-like and DUX4-rearranged subtypes in paediatric B-cell precursor acute lymphoblastic leukaemia
- PMID: 27265895
- PMCID: PMC4897744
- DOI: 10.1038/ncomms11790
Identification of ETV6-RUNX1-like and DUX4-rearranged subtypes in paediatric B-cell precursor acute lymphoblastic leukaemia
Abstract
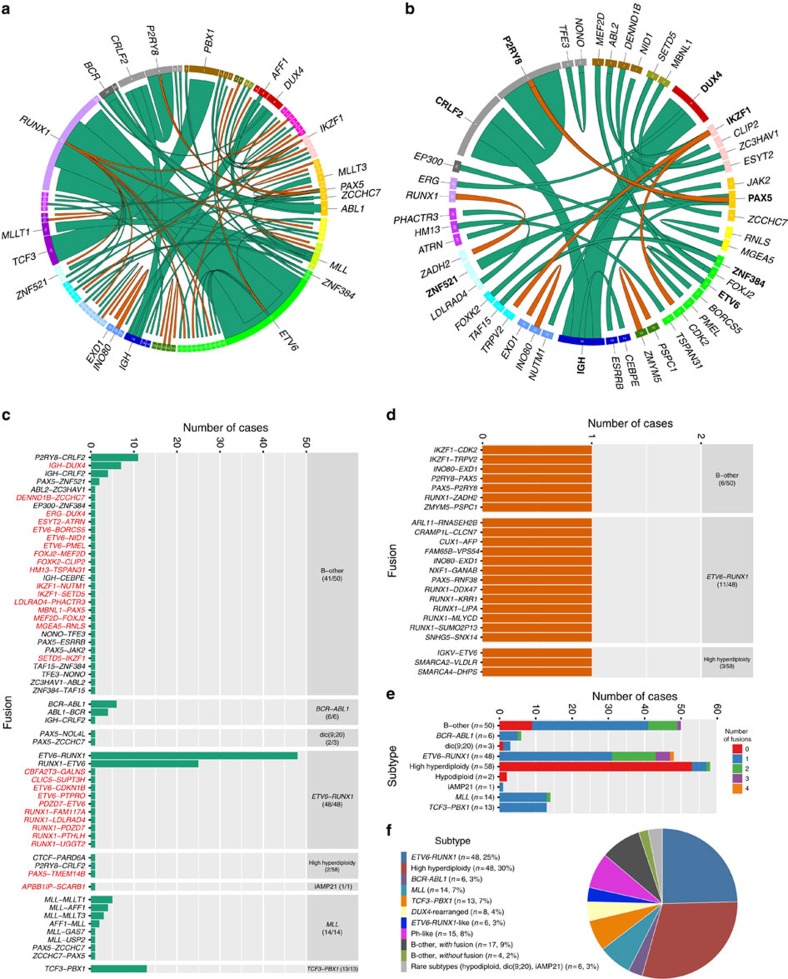
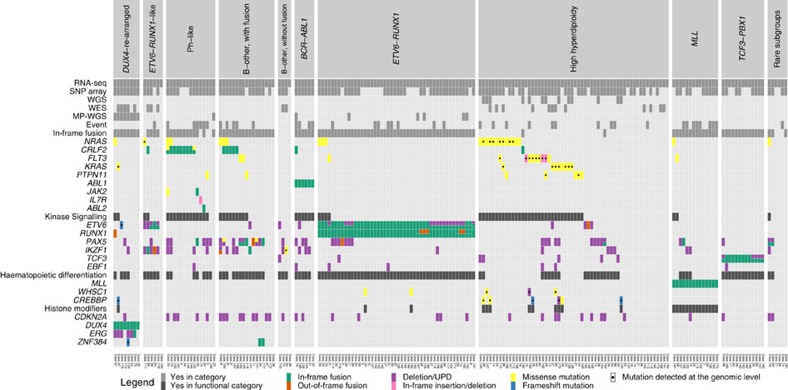
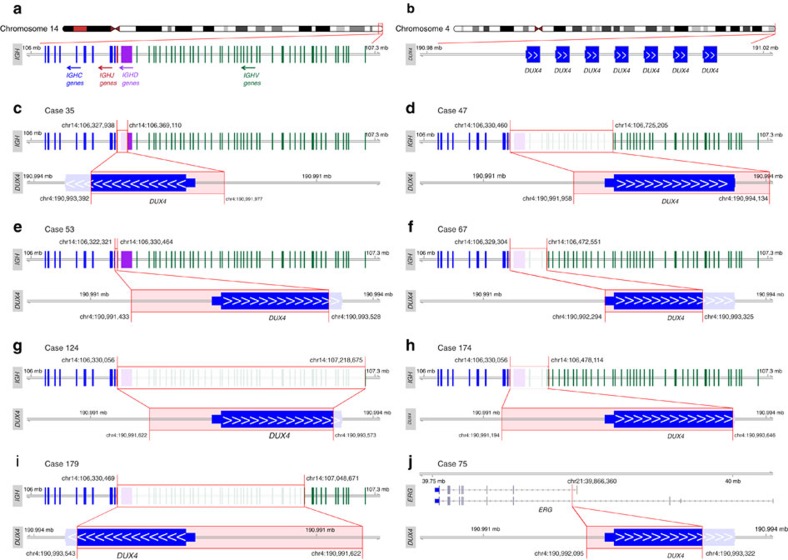
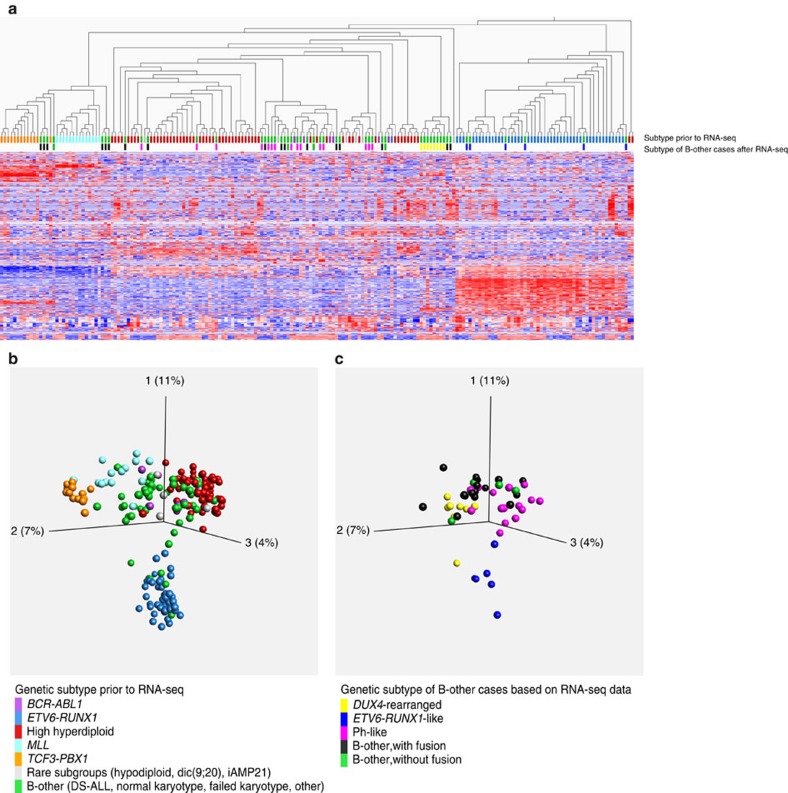
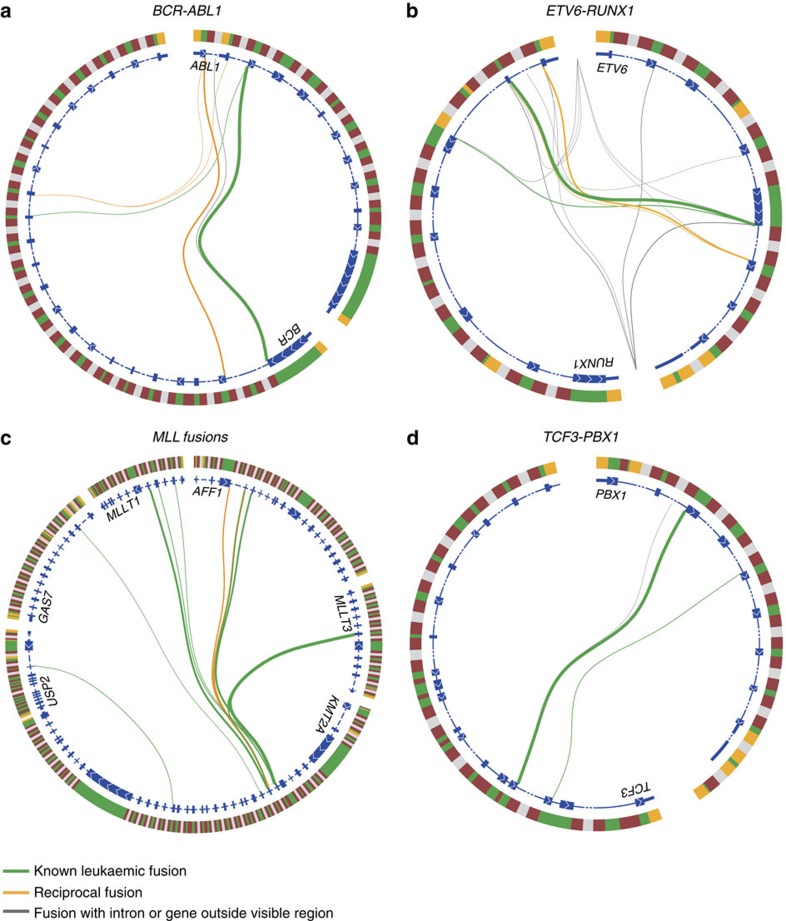
Fusion genes are potent driver mutations in cancer. In this study, we delineate the fusion gene landscape in a consecutive series of 195 paediatric B-cell precursor acute lymphoblastic leukaemia (BCP ALL). Using RNA sequencing, we find in-frame fusion genes in 127 (65%) cases, including 27 novel fusions. We describe a subtype characterized by recurrent IGH-DUX4 or ERG-DUX4 fusions, representing 4% of cases, leading to overexpression of DUX4 and frequently co-occurring with intragenic ERG deletions. Furthermore, we identify a subtype characterized by an ETV6-RUNX1-like gene-expression profile and coexisting ETV6 and IKZF1 alterations. Thus, this study provides a detailed overview of fusion genes in paediatric BCP ALL and adds new pathogenetic insights, which may improve risk stratification and provide therapeutic options for this disease.
Figures

References
-
- Pui C.-H. & Evans W. E. Treatment of acute lymphoblastic leukemia. N. Engl. J. Med. 354, 166–178 (2006). - PubMed
-
- Martinelli G. et al.. IKZF1 (Ikaros) deletions in BCR-ABL1–positive acute lymphoblastic leukemia are associated with short disease-free survival and high rate of cumulative incidence of relapse: A GIMEMA AL WP report. J. Clin. Oncol. 27, 5202–5207 (2009). - PubMed
-
- Olsson L. et al.. Deletions of IKZF1 and SPRED1 are associated with poor prognosis in a population-based series of pediatric B-cell precursor acute lymphoblastic leukemia diagnosed between 1992 and 2011. Leukemia 28, 302–310 (2014). - PubMed
