

The neural basis of depth perception from motion parallax
- PMID: 27269599
- PMCID: PMC4901450
- DOI: 10.1098/rstb.2015.0256
The neural basis of depth perception from motion parallax
Erratum in
-
Correction to 'The neural basis of depth perception from motion parallax'.Philos Trans R Soc Lond B Biol Sci. 2016 Oct 19;371(1706):20160395. doi: 10.1098/rstb.2016.0395. Philos Trans R Soc Lond B Biol Sci. 2016. PMID: 27619707 Free PMC article. No abstract available.
Abstract
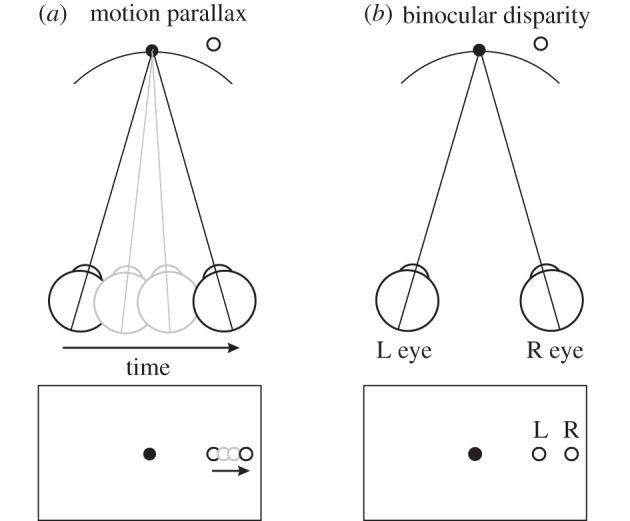
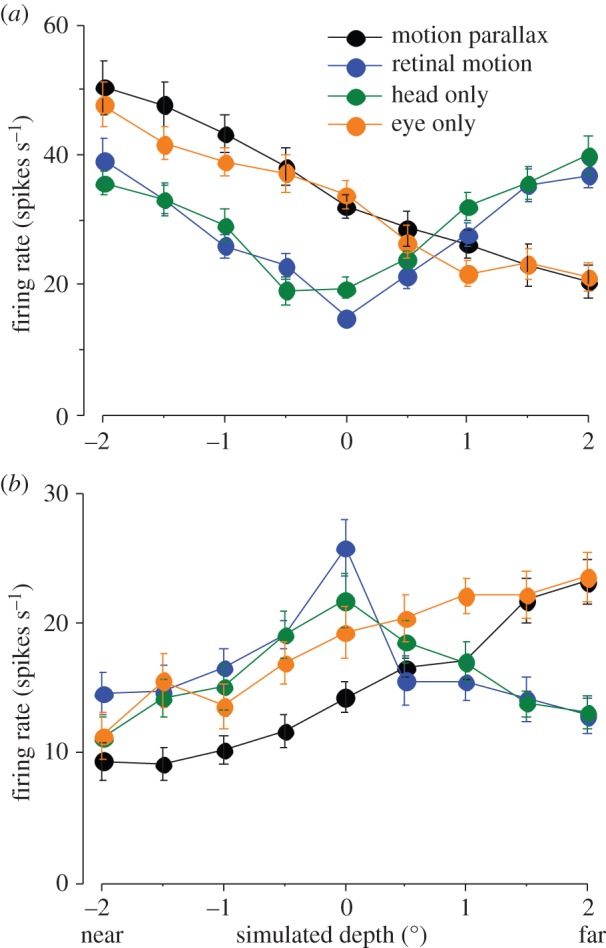
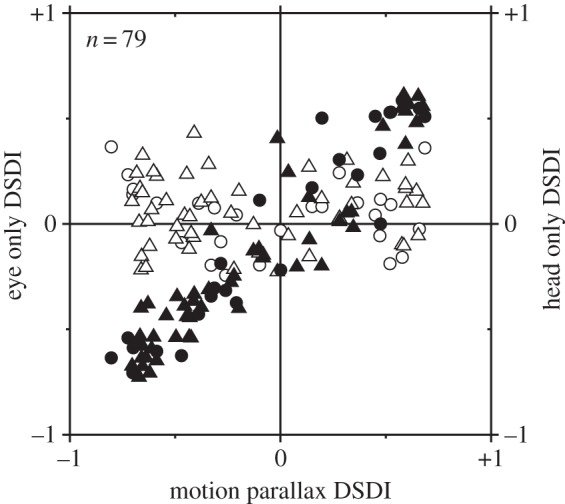
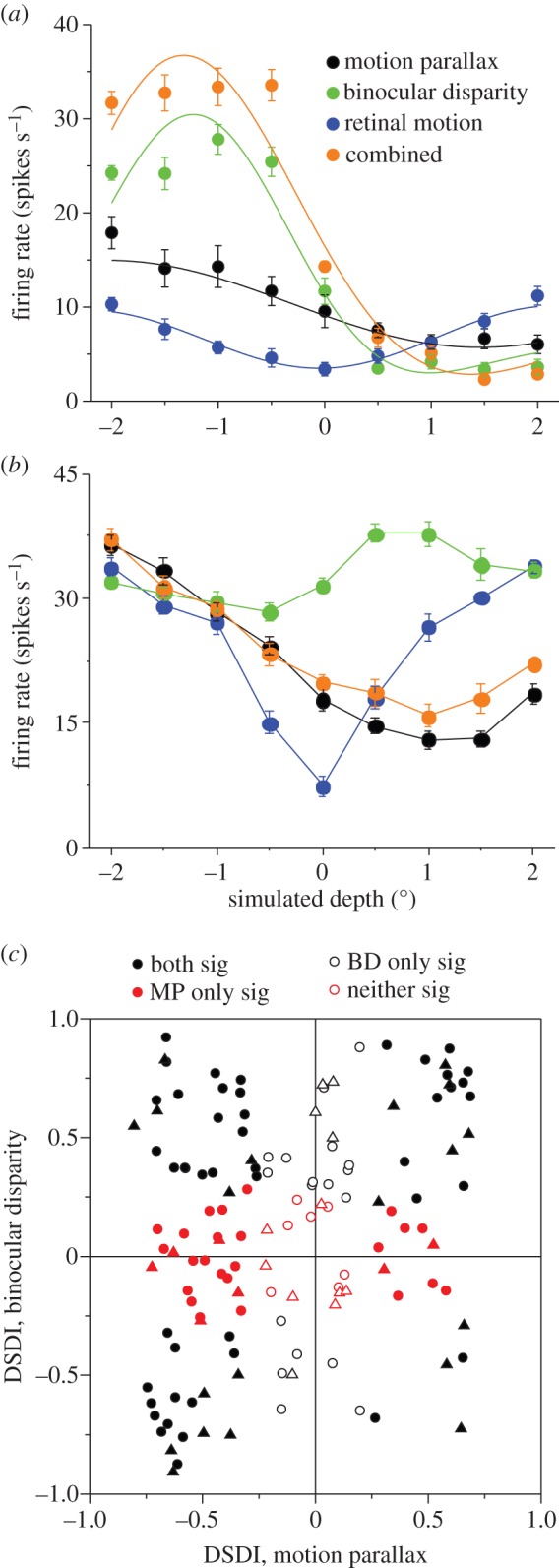
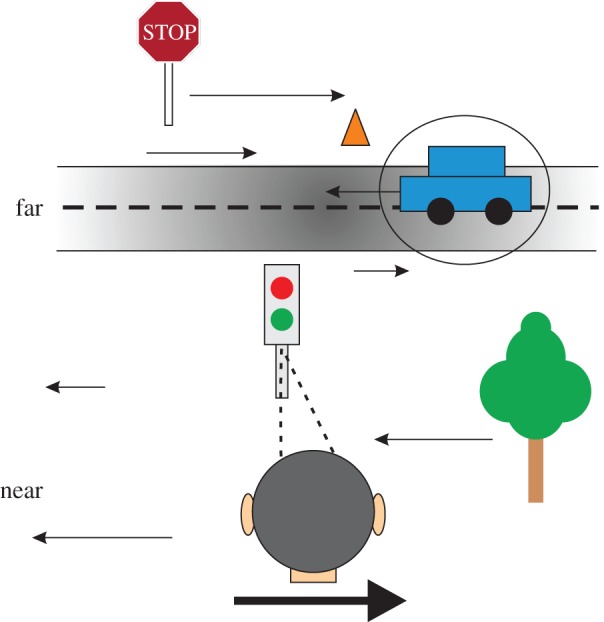
In addition to depth cues afforded by binocular vision, the brain processes relative motion signals to perceive depth. When an observer translates relative to their visual environment, the relative motion of objects at different distances (motion parallax) provides a powerful cue to three-dimensional scene structure. Although perception of depth based on motion parallax has been studied extensively in humans, relatively little is known regarding the neural basis of this visual capability. We review recent advances in elucidating the neural mechanisms for representing depth-sign (near versus far) from motion parallax. We examine a potential neural substrate in the middle temporal visual area for depth perception based on motion parallax, and we explore the nature of the signals that provide critical inputs for disambiguating depth-sign.This article is part of the themed issue 'Vision in our three-dimensional world'.
Keywords: depth; motion parallax; neural computation.
© 2016 The Author(s).
Figures



References
-
- Howard IP, Rogers BJ. 1995. Binocular vision and stereopsis. New York, NY: Oxford University Press.
-
- Howard IP, Rogers BJ. 2002. Seeing in depth. Volume 2: depth perception. Toronto, Canada: I. Porteus.
-
- Julesz B. 1971. Foundations of cylclopean perception. Chicago, IL: University of Chicago Press.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources