

Structure and calcium-binding studies of calmodulin-like domain of human non-muscle α-actinin-1
- PMID: 27272015
- PMCID: PMC4895382
- DOI: 10.1038/srep27383
Structure and calcium-binding studies of calmodulin-like domain of human non-muscle α-actinin-1
Abstract
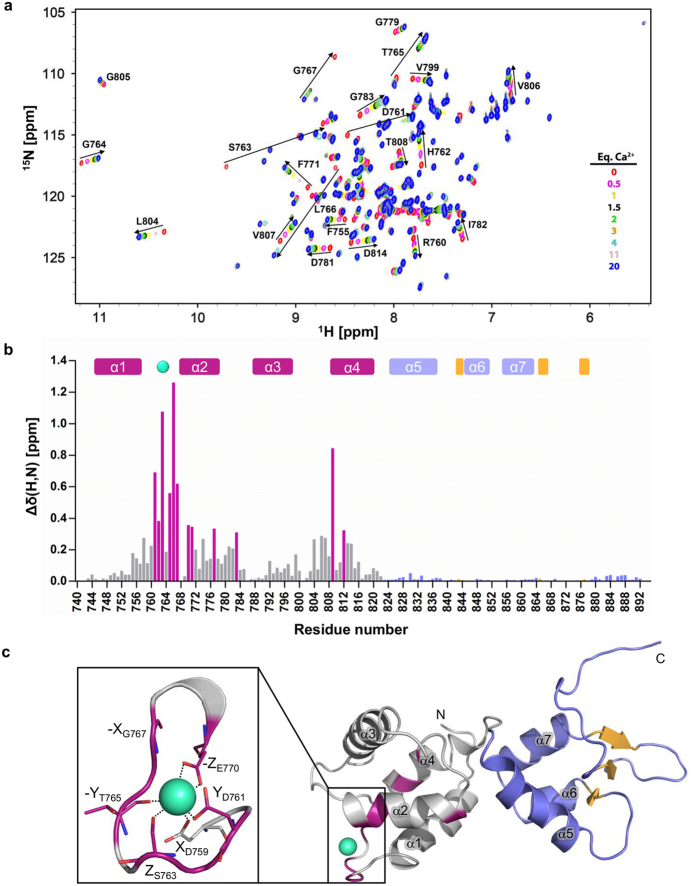
The activity of several cytosolic proteins critically depends on the concentration of calcium ions. One important intracellular calcium-sensing protein is α-actinin-1, the major actin crosslinking protein in focal adhesions and stress fibers. The actin crosslinking activity of α-actinin-1 has been proposed to be negatively regulated by calcium, but the underlying molecular mechanisms are poorly understood. To address this, we determined the first high-resolution NMR structure of its functional calmodulin-like domain (CaMD) in calcium-bound and calcium-free form. These structures reveal that in the absence of calcium, CaMD displays a conformationally flexible ensemble that undergoes a structural change upon calcium binding, leading to limited rotation of the N- and C-terminal lobes around the connecting linker and consequent stabilization of the calcium-loaded structure. Mutagenesis experiments, coupled with mass-spectrometry and isothermal calorimetry data designed to validate the calcium binding stoichiometry and binding site, showed that human non-muscle α-actinin-1 binds a single calcium ion within the N-terminal lobe. Finally, based on our structural data and analogy with other α-actinins, we provide a structural model of regulation of the actin crosslinking activity of α-actinin-1 where calcium induced structural stabilisation causes fastening of the juxtaposed actin binding domain, leading to impaired capacity to crosslink actin.
Figures







Similar articles
-
Solution structure of the calmodulin-like C-terminal domain of Entamoeba α-actinin2.Proteins. 2016 Apr;84(4):461-6. doi: 10.1002/prot.24992. Epub 2016 Feb 5. Proteins. 2016. PMID: 26800385
-
Calcium modulates calmodulin/α-actinin 1 interaction with and agonist-dependent internalization of the adenosine A2A receptor.Biochim Biophys Acta Mol Cell Res. 2017 Apr;1864(4):674-686. doi: 10.1016/j.bbamcr.2017.01.013. Epub 2017 Jan 24. Biochim Biophys Acta Mol Cell Res. 2017. PMID: 28130124
-
The three-dimensional structure of alpha-actinin obtained by cryoelectron microscopy suggests a model for Ca(2+)-dependent actin binding.J Mol Biol. 2001 Jul 20;310(4):845-58. doi: 10.1006/jmbi.2001.4789. J Mol Biol. 2001. PMID: 11453692
-
Alpha-actinin structure and regulation.Cell Mol Life Sci. 2008 Sep;65(17):2688-701. doi: 10.1007/s00018-008-8080-8. Cell Mol Life Sci. 2008. PMID: 18488141 Free PMC article. Review.
-
Proteins with calmodulin-like domains: structures and functional roles.Cell Mol Life Sci. 2019 Jun;76(12):2299-2328. doi: 10.1007/s00018-019-03062-z. Epub 2019 Mar 15. Cell Mol Life Sci. 2019. PMID: 30877334 Free PMC article. Review.
Cited by
-
Chemical shift assignments of the α-actinin C-terminal EF-hand domain bound to a cytosolic C0 domain of GluN1 (residues 841-865) from the NMDA receptor.Biomol NMR Assign. 2024 Dec;18(2):239-244. doi: 10.1007/s12104-024-10194-2. Epub 2024 Aug 29. Biomol NMR Assign. 2024. PMID: 39207574 Free PMC article.
-
Transcriptome profiling analysis of muscle tissue reveals potential candidate genes affecting water holding capacity in Chinese Simmental beef cattle.Sci Rep. 2021 Jun 7;11(1):11897. doi: 10.1038/s41598-021-91373-2. Sci Rep. 2021. PMID: 34099805 Free PMC article.
-
Calcium modulates the domain flexibility and function of an α-actinin similar to the ancestral α-actinin.Proc Natl Acad Sci U S A. 2020 Sep 8;117(36):22101-22112. doi: 10.1073/pnas.1917269117. Epub 2020 Aug 26. Proc Natl Acad Sci U S A. 2020. PMID: 32848067 Free PMC article.
-
Changes in Whey Proteome between Mediterranean and Murrah Buffalo Colostrum and Mature Milk Reflect Their Pharmaceutical and Medicinal Value.Molecules. 2022 Feb 27;27(5):1575. doi: 10.3390/molecules27051575. Molecules. 2022. PMID: 35268677 Free PMC article.
-
Getting cells into shape by calcium-dependent actin cross-linking proteins.Front Cell Dev Biol. 2023 Mar 21;11:1171930. doi: 10.3389/fcell.2023.1171930. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37025173 Free PMC article. Review.
References
-
- Djinovic-Carugo K., Gautel M., Ylanne J. & Young P. The spectrin repeat: a structural platform for cytoskeletal protein assemblies. FEBS Lett. 513, 119–123 (2002). - PubMed
-
- Otey C. A. & Carpen O. Alpha-actinin revisited: a fresh look at an old player. Cell Motil. Cytoskelet. 58, 104–111 (2004). - PubMed
-
- Foley K. S. & Young P. W. The non-muscle functions of actinins: an update. Biochem. J. 459, 1–13 (2014). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
