

Copper regulates cyclic-AMP-dependent lipolysis
- PMID: 27272565
- PMCID: PMC4955676
- DOI: 10.1038/nchembio.2098
Copper regulates cyclic-AMP-dependent lipolysis
Abstract
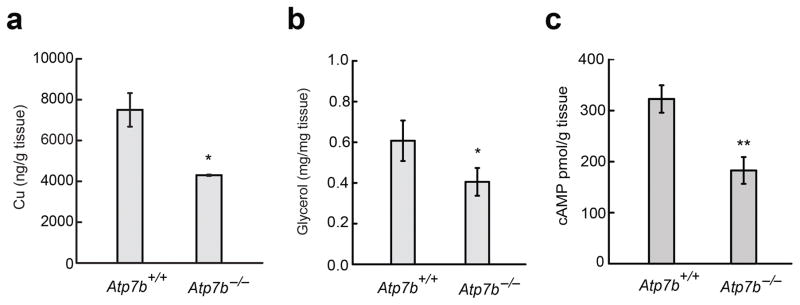
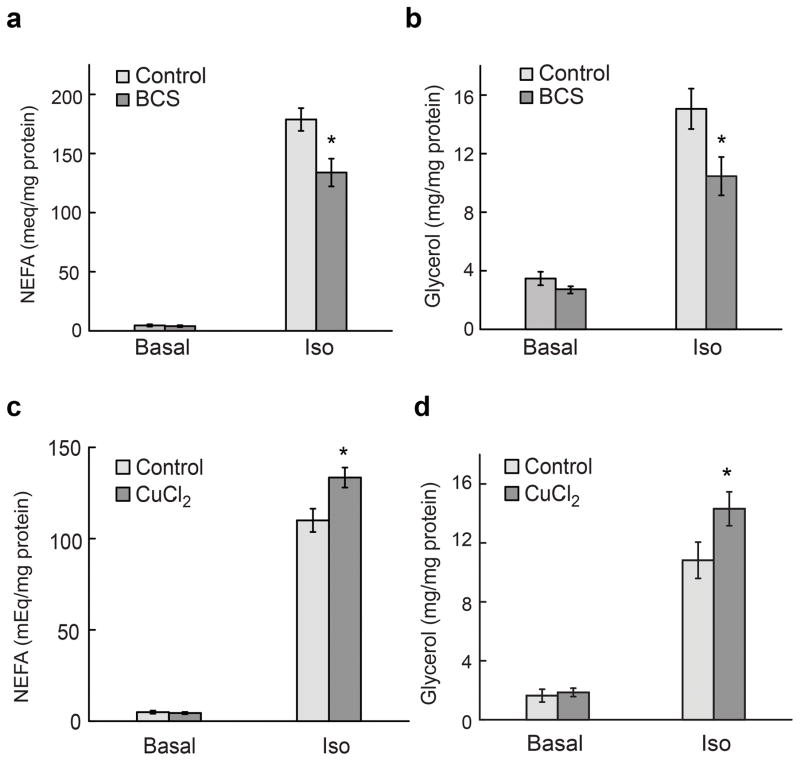
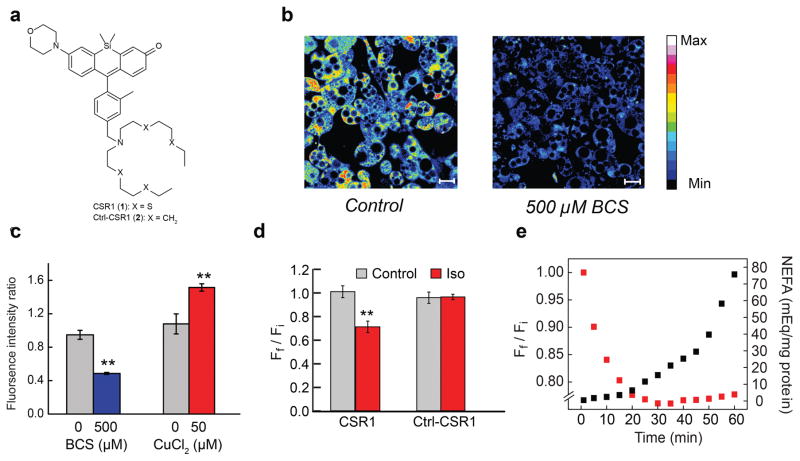
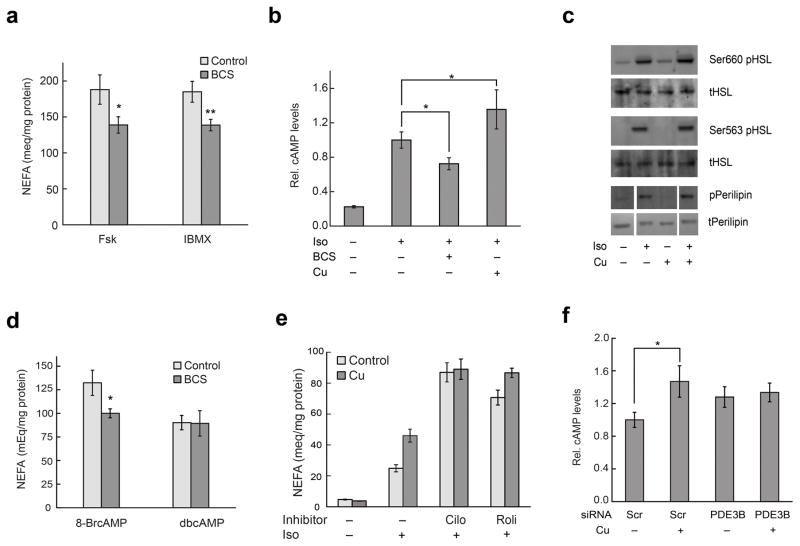
Cell signaling relies extensively on dynamic pools of redox-inactive metal ions such as sodium, potassium, calcium and zinc, but their redox-active transition metal counterparts such as copper and iron have been studied primarily as static enzyme cofactors. Here we report that copper is an endogenous regulator of lipolysis, the breakdown of fat, which is an essential process in maintaining body weight and energy stores. Using a mouse model of genetic copper misregulation, in combination with pharmacological alterations in copper status and imaging studies in a 3T3-L1 white adipocyte model, we found that copper regulates lipolysis at the level of the second messenger, cyclic AMP (cAMP), by altering the activity of the cAMP-degrading phosphodiesterase PDE3B. Biochemical studies of the copper-PDE3B interaction establish copper-dependent inhibition of enzyme activity and identify a key conserved cysteine residue in a PDE3-specific loop that is essential for the observed copper-dependent lipolytic phenotype.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

References
-
- Lippard SJ, Berg JM. Principles of Bioinorganic Chemistry. University Science Books; Mill Valley, CA: 1994.
-
- Barnham KJ, Masters CL, Bush AI. Neurodegenerative diseases and oxidative stress. Nat Rev Drug Discov. 2004;3:205–14. - PubMed
-
- Rae TD, Schmidt PJ, Pufahl RA, Culotta VC, O'Halloran TV. Undetectable intracellular free copper: the requirement of a copper chaperone for superoxide dismutase. Science. 1999;284:805–8. - PubMed
-
- Banci L, et al. Affinity gradients drive copper to cellular destinations. Nature. 2010;465:645–8. - PubMed
Publication types
MeSH terms
Substances
Associated data
- PubChem-Substance/313181569
- PubChem-Substance/313181570
- PubChem-Substance/313181571
- PubChem-Substance/313181572
- PubChem-Substance/313181573
- PubChem-Substance/313181574
- PubChem-Substance/313181575
- PubChem-Substance/313181576
- PubChem-Substance/313181577
- PubChem-Substance/313181578
- PubChem-Substance/313181579
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
