

Plasma membrane/cell wall perturbation activates a novel cell cycle checkpoint during G1 in Saccharomyces cerevisiae
- PMID: 27274080
- PMCID: PMC4922162
- DOI: 10.1073/pnas.1523824113
Plasma membrane/cell wall perturbation activates a novel cell cycle checkpoint during G1 in Saccharomyces cerevisiae
Abstract
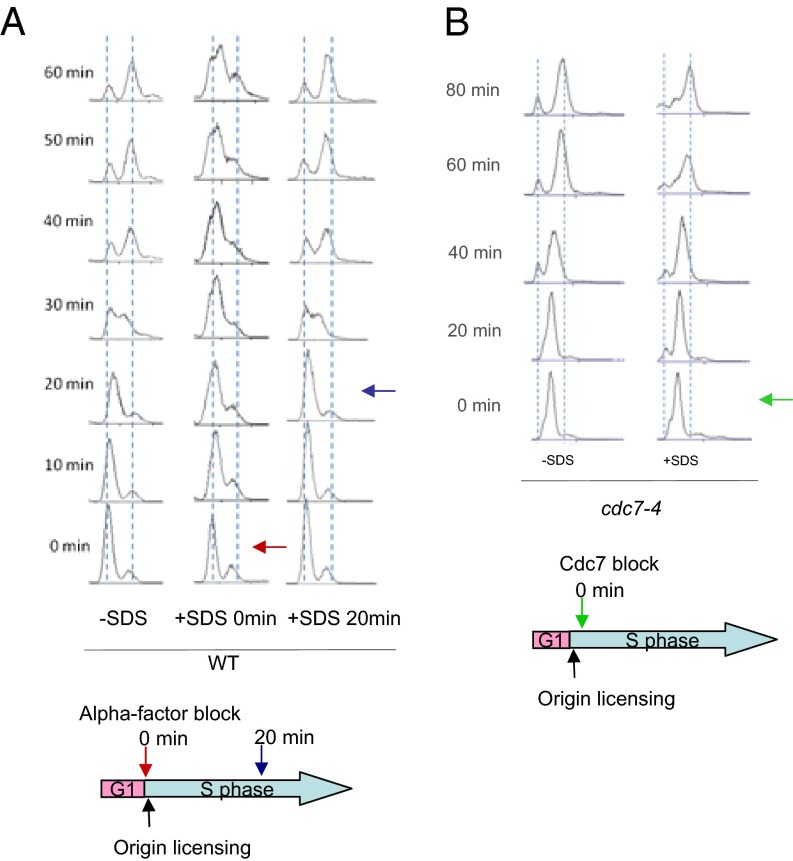
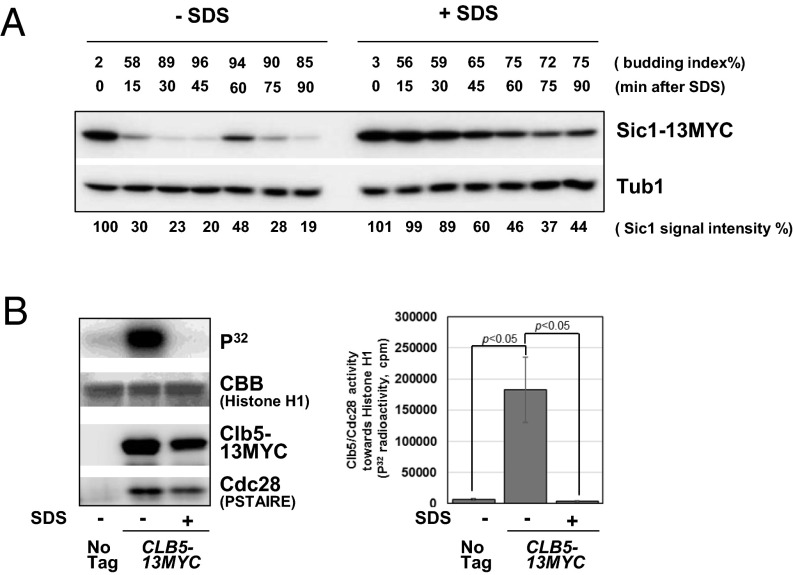
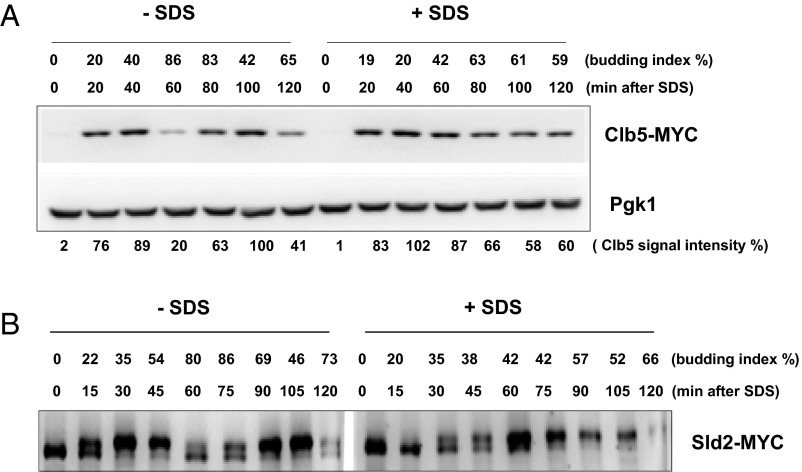
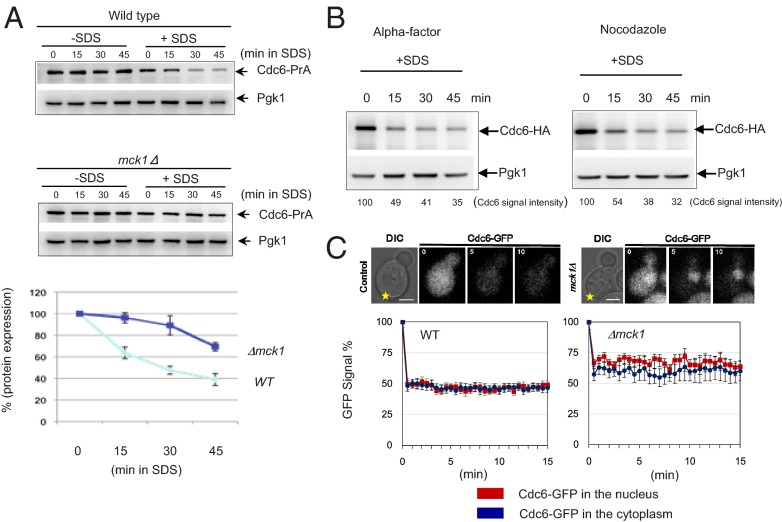
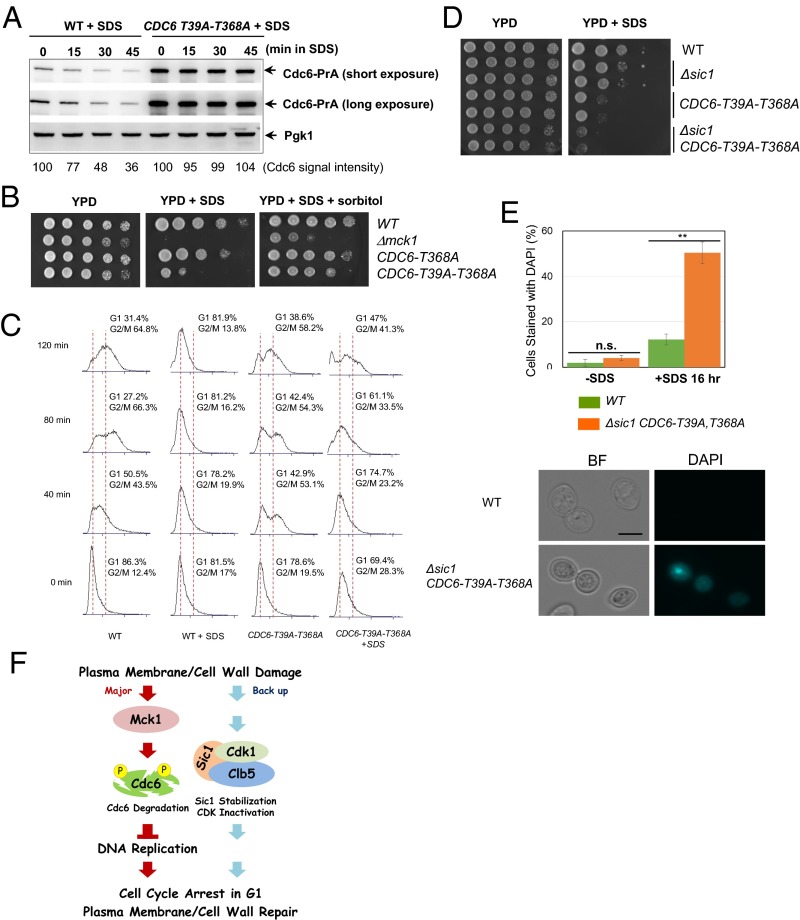
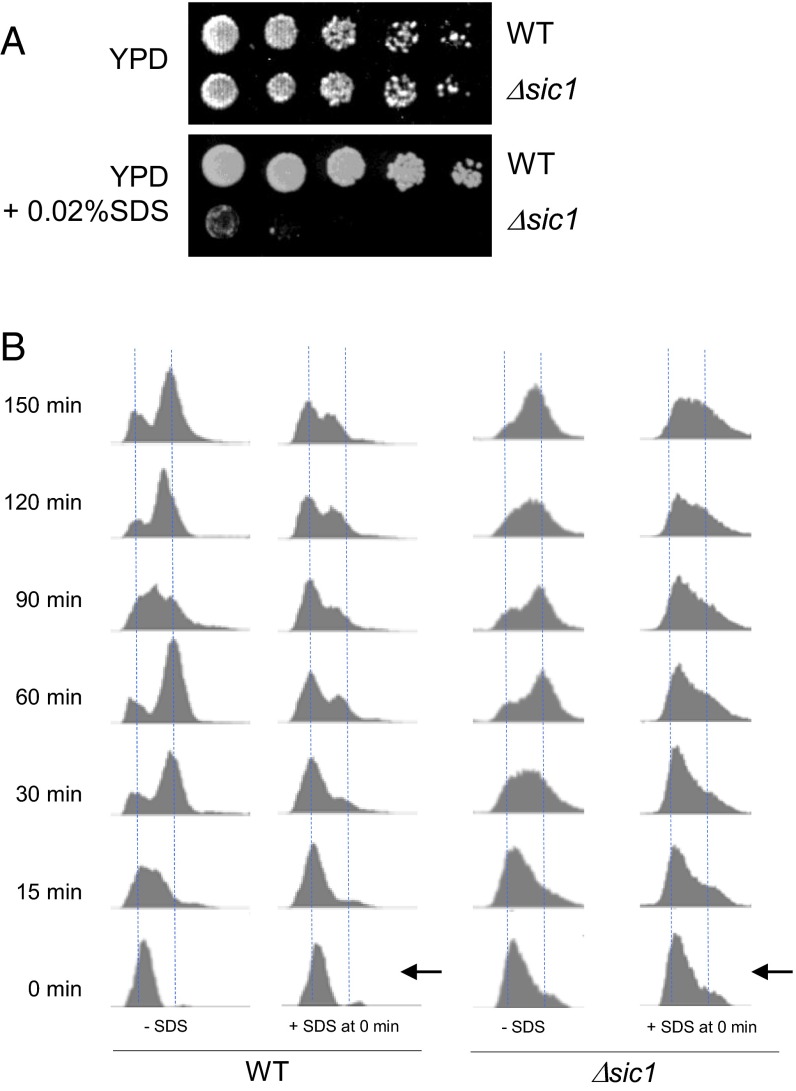
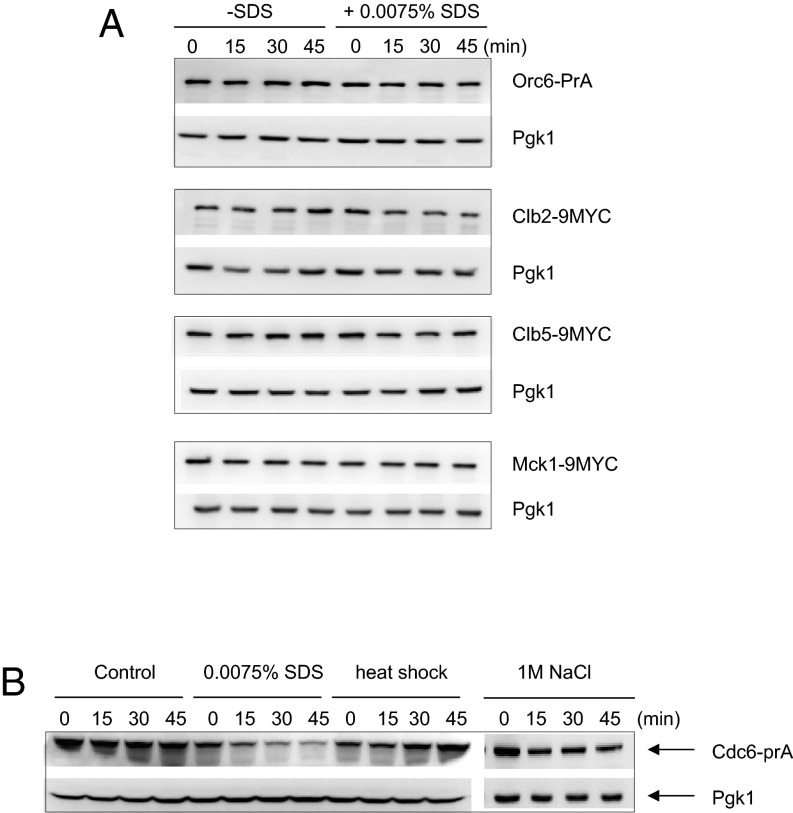
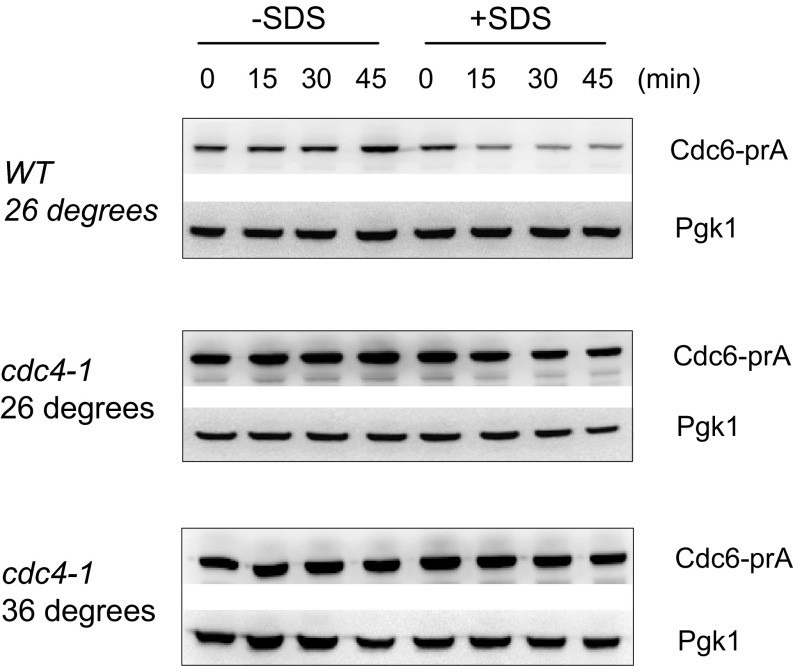
Cellular wound healing or the repair of plasma membrane/cell wall damage (plasma membrane damage) occurs frequently in nature. Although various cellular perturbations, such as DNA damage, spindle misalignment, and impaired daughter cell formation, are monitored by cell cycle checkpoint mechanisms in budding yeast, whether plasma membrane damage is monitored by any of these checkpoints remains to be addressed. Here, we define the mechanism by which cells sense membrane damage and inhibit DNA replication. We found that the inhibition of DNA replication upon plasma membrane damage requires GSK3/Mck1-dependent degradation of Cdc6, a component of the prereplicative complex. Furthermore, the CDK inhibitor Sic1 is stabilized in response to plasma membrane damage, leading to cell integrity maintenance in parallel with the Mck1-Cdc6 pathway. Cells defective in both Cdc6 degradation and Sic1 stabilization failed to grow in the presence of plasma membrane damage. Taking these data together, we propose that plasma membrane damage triggers G1 arrest via Cdc6 degradation and Sic1 stabilization to promote the cellular wound healing process.
Keywords: DNA replication; cdc6; cell wall integrity; mck1; sic1.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Cdc6 cooperates with Sic1 and Hct1 to inactivate mitotic cyclin-dependent kinases.Nature. 2001 Jul 19;412(6844):355-8. doi: 10.1038/35085610. Nature. 2001. PMID: 11460169
-
CDK Pho85 targets CDK inhibitor Sic1 to relieve yeast G1 checkpoint arrest after DNA damage.Nat Struct Mol Biol. 2006 Oct;13(10):908-14. doi: 10.1038/nsmb1139. Epub 2006 Sep 10. Nat Struct Mol Biol. 2006. PMID: 16964260
-
Cdc6 degradation requires phosphodegron created by GSK-3 and Cdk1 for SCFCdc4 recognition in Saccharomyces cerevisiae.Mol Biol Cell. 2015 Jul 15;26(14):2609-19. doi: 10.1091/mbc.E14-07-1213. Epub 2015 May 20. Mol Biol Cell. 2015. PMID: 25995377 Free PMC article.
-
Mechanisms involved in regulating DNA replication origins during the cell cycle and in response to DNA damage.Philos Trans R Soc Lond B Biol Sci. 2004 Jan 29;359(1441):31-8. doi: 10.1098/rstb.2003.1362. Philos Trans R Soc Lond B Biol Sci. 2004. PMID: 15065654 Free PMC article. Review.
-
G1 regulation and checkpoints operating around START in fission yeast.Bioessays. 1995 Jun;17(6):481-90. doi: 10.1002/bies.950170604. Bioessays. 1995. PMID: 7575489 Review.
Cited by
-
Tryptophan confers resistance to SDS-associated cell membrane stress in Saccharomyces cerevisiae.PLoS One. 2019 Mar 11;14(3):e0199484. doi: 10.1371/journal.pone.0199484. eCollection 2019. PLoS One. 2019. PMID: 30856175 Free PMC article.
-
The Role of Mechanoperception in Plant Cell Wall Integrity Maintenance.Plants (Basel). 2020 May 1;9(5):574. doi: 10.3390/plants9050574. Plants (Basel). 2020. PMID: 32369932 Free PMC article. Review.
-
Balancing of the mitotic exit network and cell wall integrity signaling governs the development and pathogenicity in Magnaporthe oryzae.PLoS Pathog. 2021 Jan 7;17(1):e1009080. doi: 10.1371/journal.ppat.1009080. eCollection 2021 Jan. PLoS Pathog. 2021. PMID: 33411855 Free PMC article.
-
Loss of tolerance to multiple environmental stresses due to limitation of structural diversity of complex sphingolipids.Mol Biol Cell. 2022 Oct 1;33(12):ar105. doi: 10.1091/mbc.E22-04-0117. Epub 2022 Jul 27. Mol Biol Cell. 2022. PMID: 35895092 Free PMC article.
-
M-O-M mediated denaturation resistant P2 tetramer on the infected erythrocyte surface of malaria parasite imports serum fatty acids.iScience. 2024 Apr 16;27(5):109760. doi: 10.1016/j.isci.2024.109760. eCollection 2024 May 17. iScience. 2024. PMID: 38726364 Free PMC article.
References
-
- Levin DE, Fields FO, Kunisawa R, Bishop JM, Thorner J. A candidate protein kinase C gene, PKC1, is required for the S. cerevisiae cell cycle. Cell. 1990;62(2):213–224. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
