

Environmental changes define ecological limits to species richness and reveal the mode of macroevolutionary competition
- PMID: 27278857
- PMCID: PMC4999050
- DOI: 10.1111/ele.12626
Environmental changes define ecological limits to species richness and reveal the mode of macroevolutionary competition
Erratum in
-
Corrigendum for Ezard and Purvis (2016) DOI: 10.1111/ele.12626.Ecol Lett. 2017 Nov;20(11):1491. doi: 10.1111/ele.12855. Ecol Lett. 2017. PMID: 29044972 Free PMC article. No abstract available.
Abstract
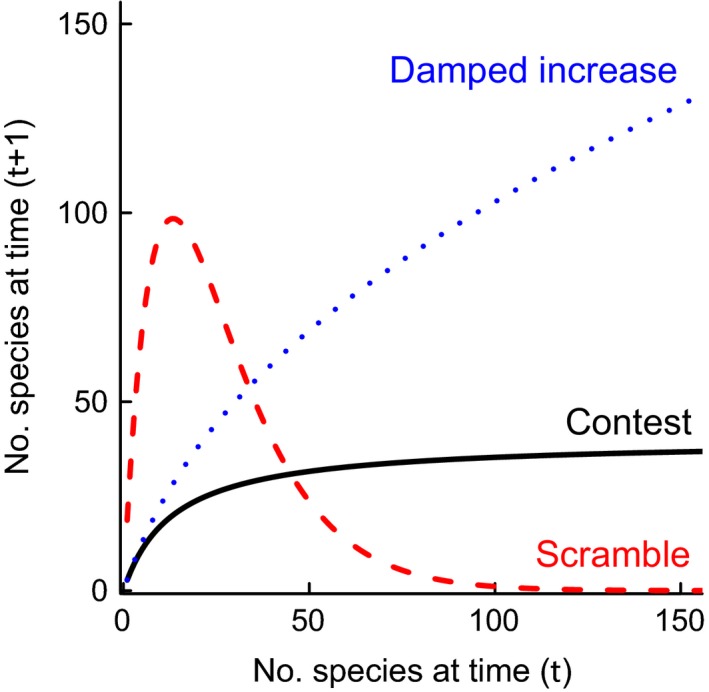
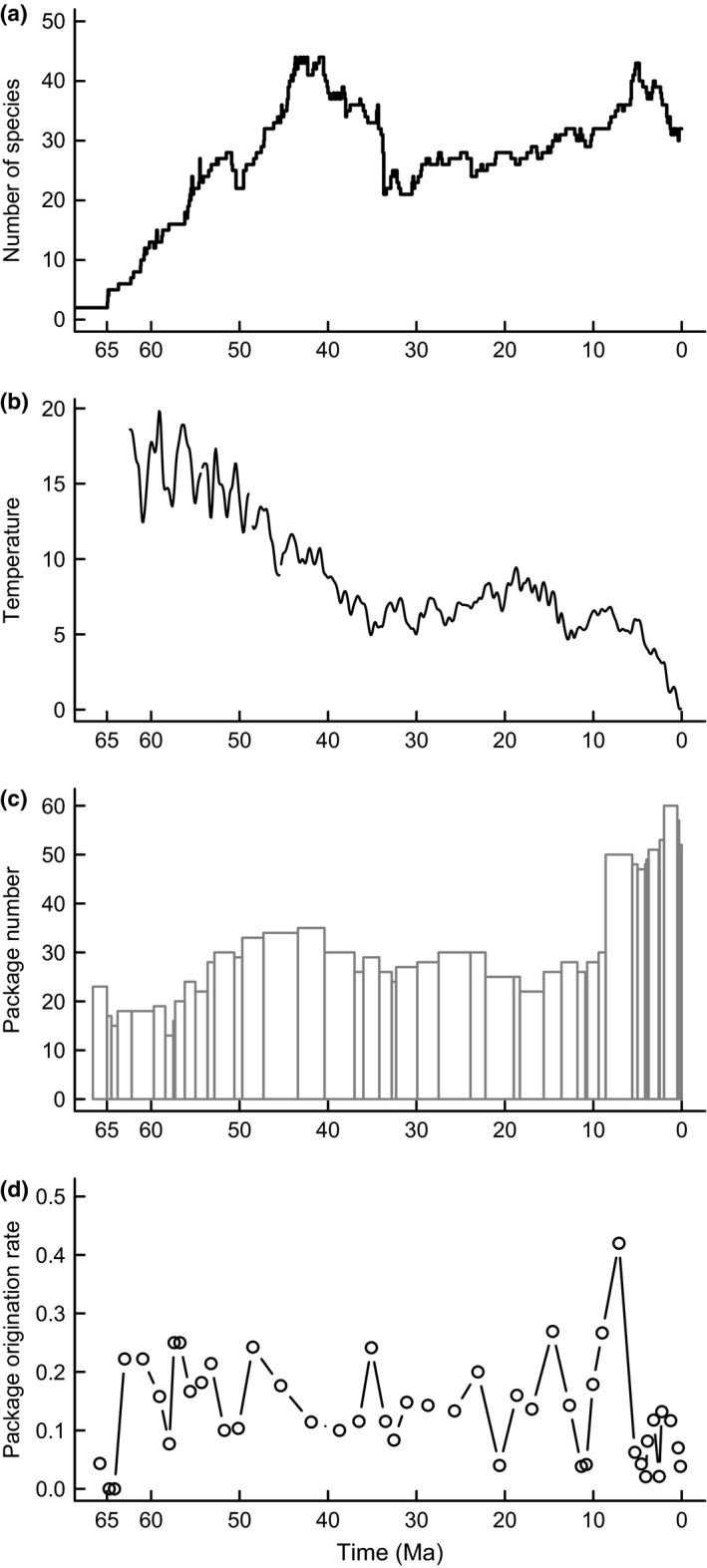
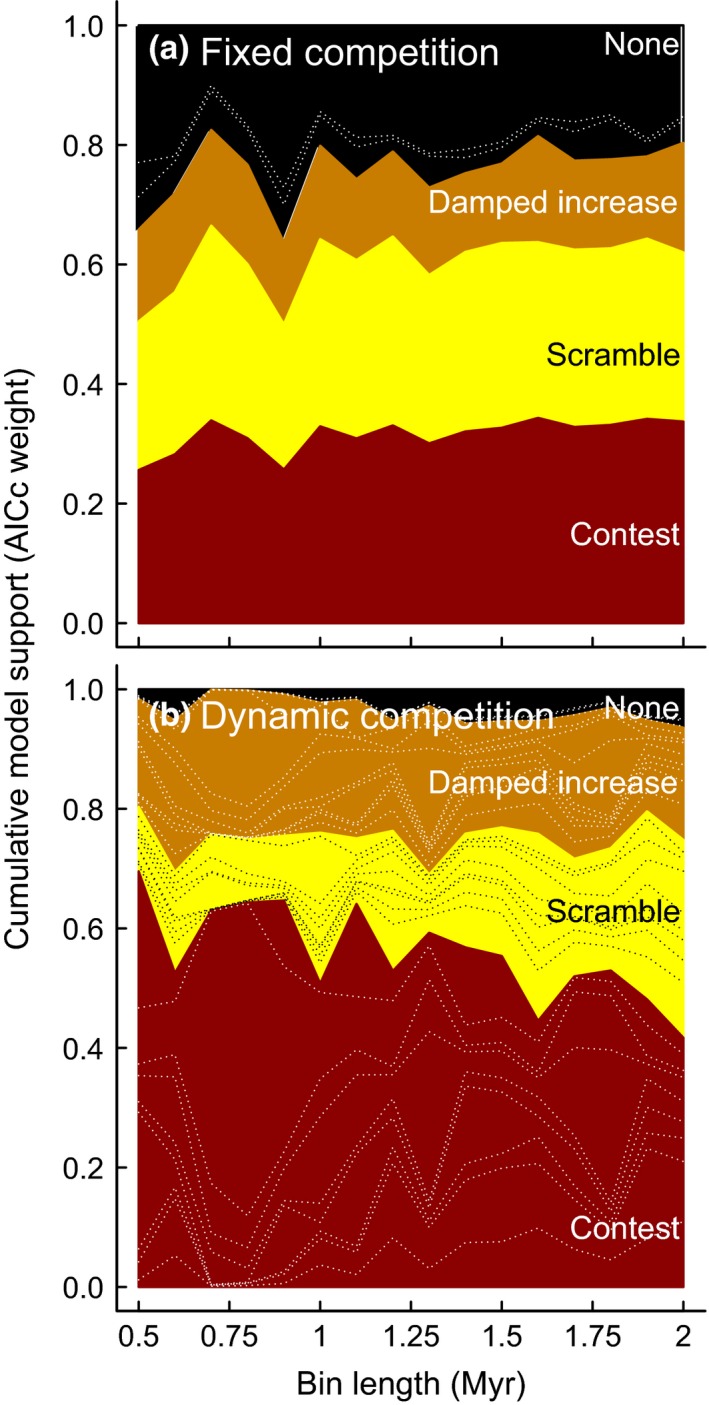
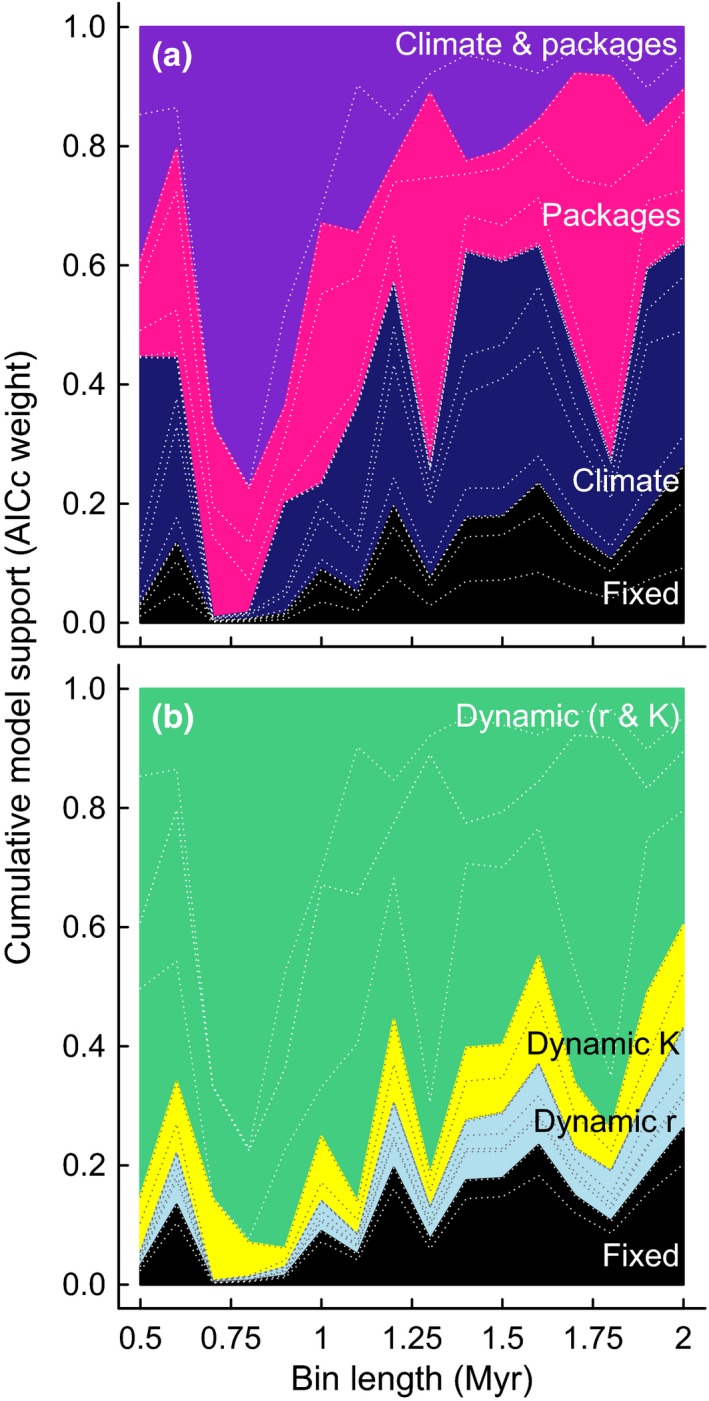
Co-dependent geological and climatic changes obscure how species interact in deep time. The interplay between these environmental factors makes it hard to discern whether ecological competition exerts an upper limit on species richness. Here, using the exceptional fossil record of Cenozoic Era macroperforate planktonic foraminifera, we assess the evidence for alternative modes of macroevolutionary competition. Our models support an environmentally dependent macroevolutionary form of contest competition that yields finite upper bounds on species richness. Models of biotic competition assuming unchanging environmental conditions were overwhelmingly rejected. In the best-supported model, temperature affects the per-lineage diversification rate, while both temperature and an environmental driver of sediment accumulation defines the upper limit. The support for contest competition implies that incumbency constrains species richness by restricting niche availability, and that the number of macroevolutionary niches varies as a function of environmental changes.
Keywords: Beverton-Holt; Ricker; contest competition; diversification; diversity-dependence; ecological limits; microfossil; scramble competition.
© 2016 The Authors. Ecology Letters published by CNRS and John Wiley & Sons Ltd.
Figures
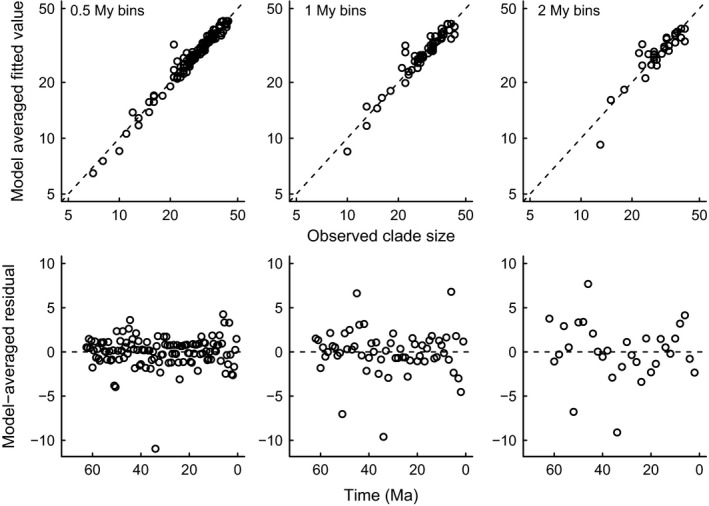
References
-
- Alroy, J. (1996). Constant extinction, constrained diversification, and uncoordinated stasis in North American mammals. Palaeogeog., Palaeoclimatol., Palaeoecol., 127, 285–311.
-
- Alroy, J. (2010). The shifting balance of diversity among major marine animal groups. Science, 329, 1191–1194. - PubMed
-
- Aze, T. , Ezard, T.H.G. , Purvis, A. , Coxall, H.K. , Stewart, D.R.M. , Wade, B.S. et al (2011). A phylogeny of Cenozoic macroperforate planktonic foraminifera from fossil data. Biol. Rev., 86, 900–927. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
