

Homology-Based Identification of a Mutation in the Coronavirus RNA-Dependent RNA Polymerase That Confers Resistance to Multiple Mutagens
- PMID: 27279608
- PMCID: PMC4984655
- DOI: 10.1128/JVI.00080-16
Homology-Based Identification of a Mutation in the Coronavirus RNA-Dependent RNA Polymerase That Confers Resistance to Multiple Mutagens
Abstract
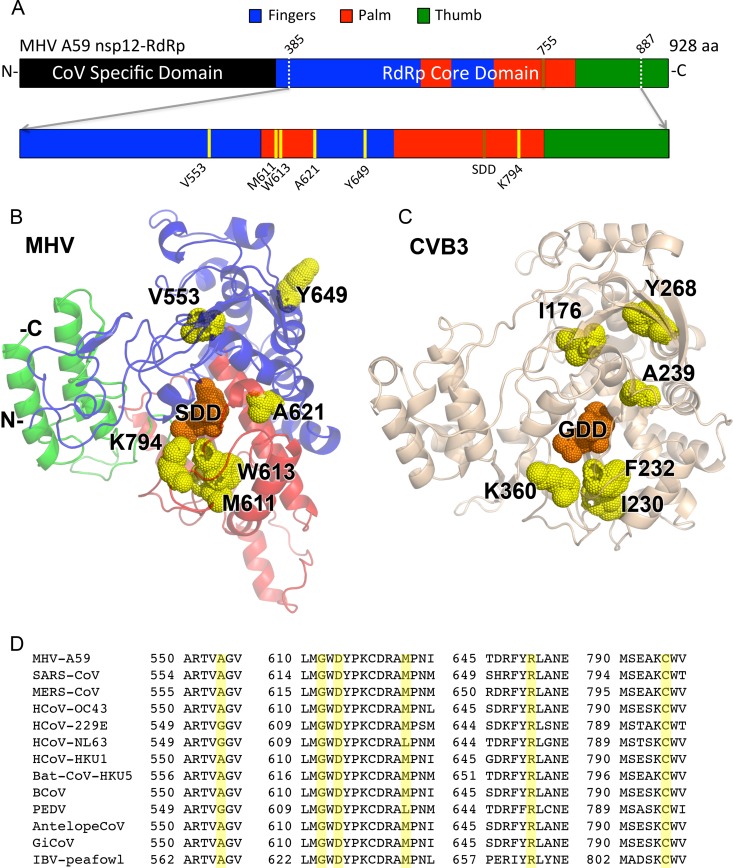
Positive-sense RNA viruses encode RNA-dependent RNA polymerases (RdRps) essential for genomic replication. With the exception of the large nidoviruses, such as coronaviruses (CoVs), RNA viruses lack proofreading and thus are dependent on RdRps to control nucleotide selectivity and fidelity. CoVs encode a proofreading exonuclease in nonstructural protein 14 (nsp14-ExoN), which confers a greater-than-10-fold increase in fidelity compared to other RNA viruses. It is unknown to what extent the CoV polymerase (nsp12-RdRp) participates in replication fidelity. We sought to determine whether homology modeling could identify putative determinants of nucleotide selectivity and fidelity in CoV RdRps. We modeled the CoV murine hepatitis virus (MHV) nsp12-RdRp structure and superimposed it on solved picornaviral RdRp structures. Fidelity-altering mutations previously identified in coxsackie virus B3 (CVB3) were mapped onto the nsp12-RdRp model structure and then engineered into the MHV genome with [nsp14-ExoN(+)] or without [nsp14-ExoN(-)] ExoN activity. Using this method, we identified two mutations conferring resistance to the mutagen 5-fluorouracil (5-FU): nsp12-M611F and nsp12-V553I. For nsp12-V553I, we also demonstrate resistance to the mutagen 5-azacytidine (5-AZC) and decreased accumulation of mutations. Resistance to 5-FU, and a decreased number of genomic mutations, was effectively masked by nsp14-ExoN proofreading activity. These results indicate that nsp12-RdRp likely functions in fidelity regulation and that, despite low sequence conservation, some determinants of RdRp nucleotide selectivity are conserved across RNA viruses. The results also indicate that, with regard to nucleotide selectivity, nsp14-ExoN is epistatic to nsp12-RdRp, consistent with its proposed role in a multiprotein replicase-proofreading complex.
Importance: RNA viruses have evolutionarily fine-tuned replication fidelity to balance requirements for genetic stability and diversity. Responsibility for replication fidelity in RNA viruses has been attributed to the RNA-dependent RNA polymerases, with mutations in RdRps for multiple RNA viruses shown to alter fidelity and attenuate virus replication and virulence. Coronaviruses (CoVs) are the only known RNA viruses to encode a proofreading exonuclease (nsp14-ExoN), as well as other replicase proteins involved in regulation of fidelity. This report shows that the CoV RdRp (nsp12) likely functions in replication fidelity; that residue determinants of CoV RdRp nucleotide selectivity map to similar structural regions of other, unrelated RNA viral polymerases; and that for CoVs, the proofreading activity of the nsp14-ExoN is epistatic to the function of the RdRp in fidelity.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Schaaper RM. 1993. Base selection, proofreading, and mismatch repair during DNA replication in Escherichia coli. J Biol Chem 268:23762–23765. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
