

α-Synuclein binds to TOM20 and inhibits mitochondrial protein import in Parkinson's disease
- PMID: 27280685
- PMCID: PMC5016095
- DOI: 10.1126/scitranslmed.aaf3634
α-Synuclein binds to TOM20 and inhibits mitochondrial protein import in Parkinson's disease
Abstract
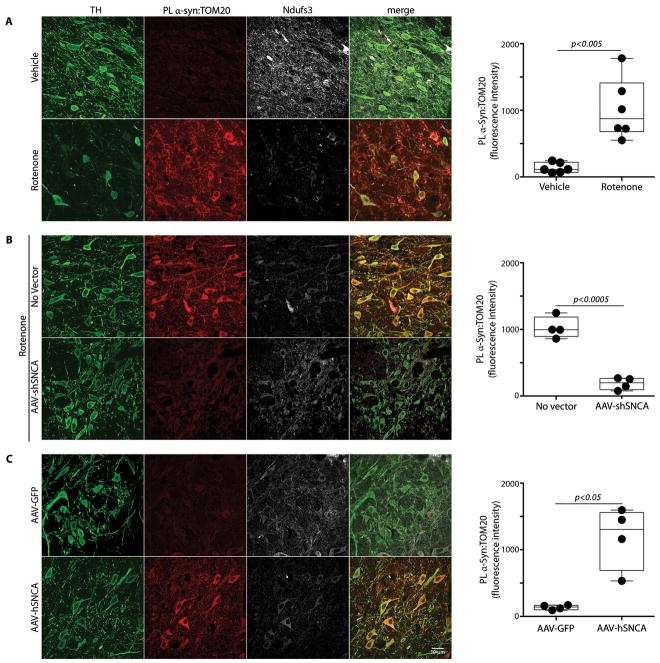
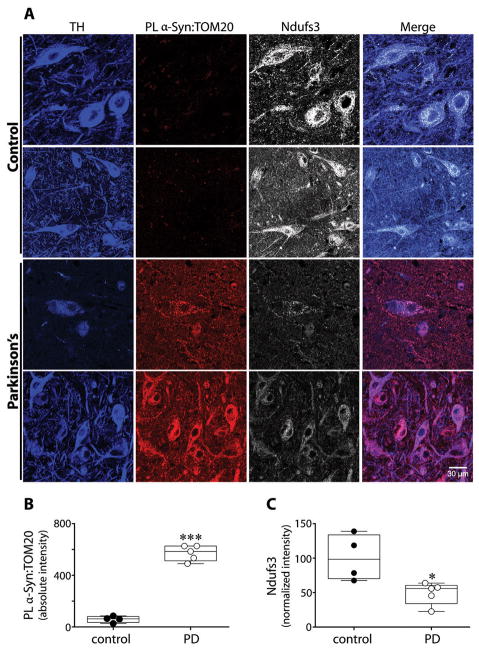
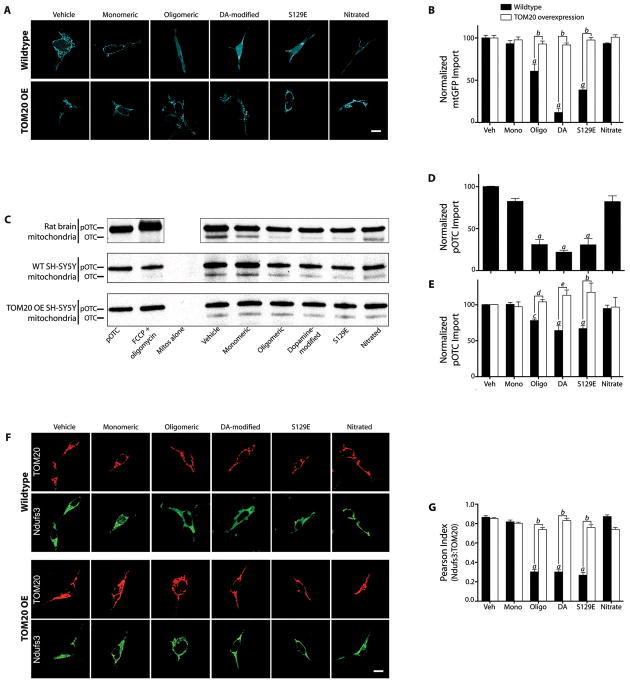
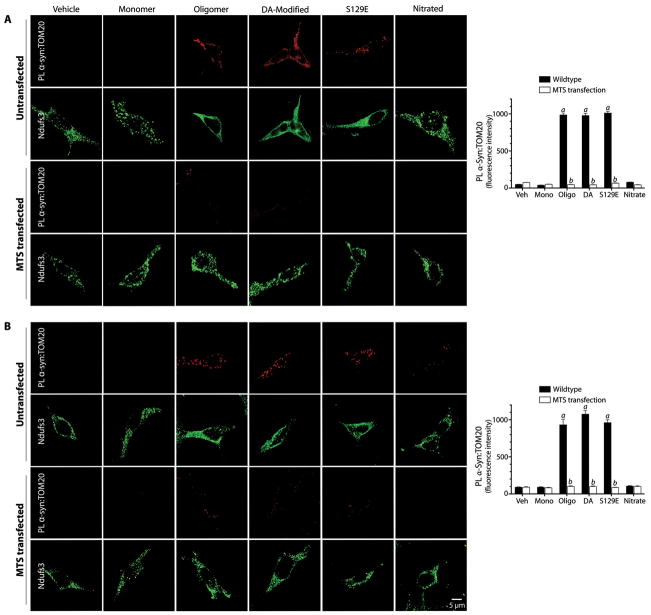
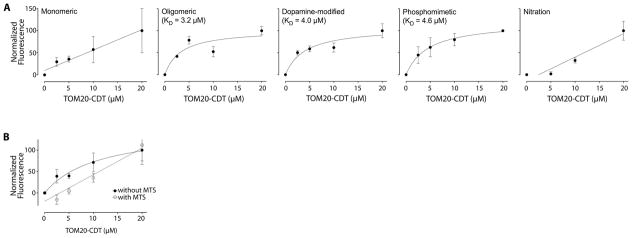
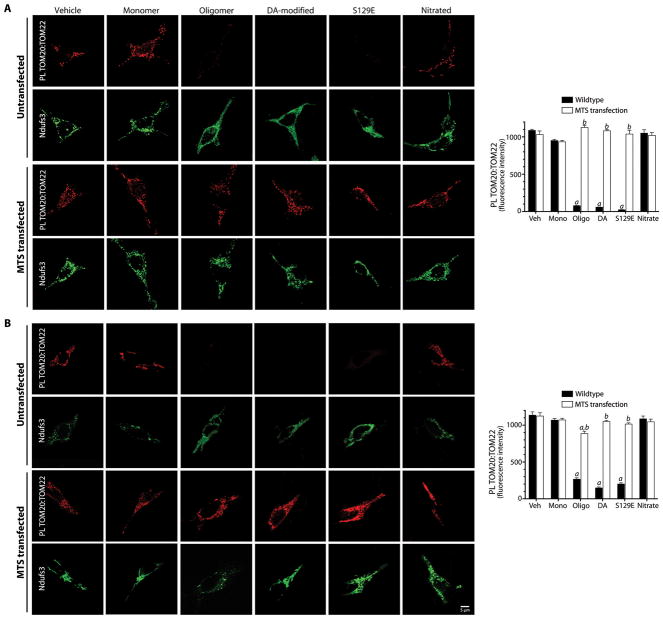
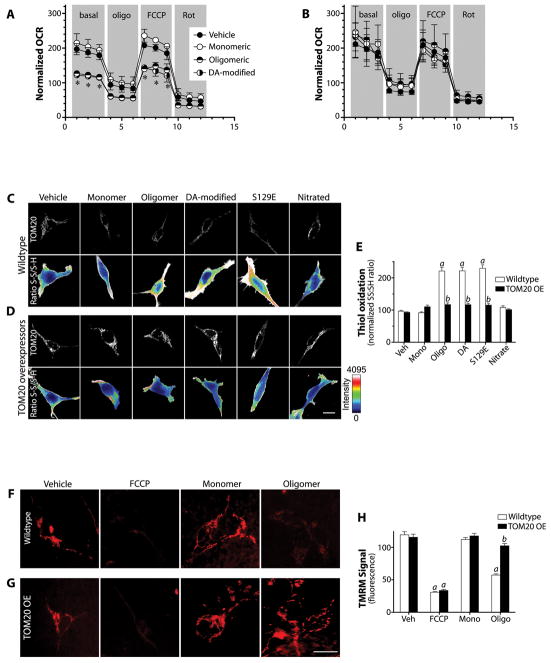
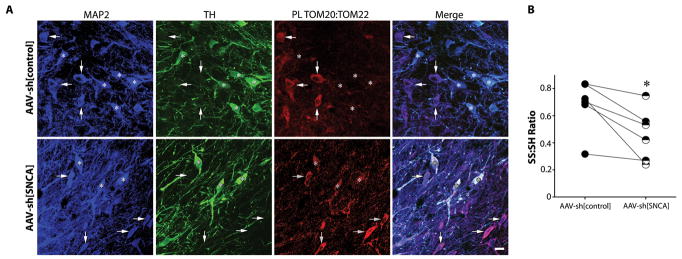
α-Synuclein accumulation and mitochondrial dysfunction have both been strongly implicated in the pathogenesis of Parkinson's disease (PD), and the two appear to be related. Mitochondrial dysfunction leads to accumulation and oligomerization of α-synuclein, and increased levels of α-synuclein cause mitochondrial impairment, but the basis for this bidirectional interaction remains obscure. We now report that certain posttranslationally modified species of α-synuclein bind with high affinity to the TOM20 (translocase of the outer membrane 20) presequence receptor of the mitochondrial protein import machinery. This binding prevented the interaction of TOM20 with its co-receptor, TOM22, and impaired mitochondrial protein import. Consequently, there were deficient mitochondrial respiration, enhanced production of reactive oxygen species, and loss of mitochondrial membrane potential. Examination of postmortem brain tissue from PD patients revealed an aberrant α-synuclein-TOM20 interaction in nigrostriatal dopaminergic neurons that was associated with loss of imported mitochondrial proteins, thereby confirming this pathogenic process in the human disease. Modest knockdown of endogenous α-synuclein was sufficient to maintain mitochondrial protein import in an in vivo model of PD. Furthermore, in in vitro systems, overexpression of TOM20 or a mitochondrial targeting signal peptide had beneficial effects and preserved mitochondrial protein import. This study characterizes a pathogenic mechanism in PD, identifies toxic species of wild-type α-synuclein, and reveals potential new therapeutic strategies for neuroprotection.
Copyright © 2016, American Association for the Advancement of Science.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures
Comment in
-
New evidence comes to light: How is α-synuclein aggregation related to mitochondrial protein import in Parkinson's disease?Mov Disord. 2017 Jan;32(1):107. doi: 10.1002/mds.26889. Epub 2016 Dec 22. Mov Disord. 2017. PMID: 28004858 No abstract available.
References
-
- Kalia LV, Lang AE. Parkinson’s disease. Lancet. 2015 Apr 17; - PubMed
-
- Betarbet R, et al. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat Neurosci. 2000 Dec;3:1301. - PubMed
-
- Betarbet R, et al. Intersecting pathways to neurodegeneration in Parkinson’s disease: effects of the pesticide rotenone on DJ-1, alpha-synuclein, and the ubiquitin-proteasome system. Neurobiol Dis. 2006 May;22:404. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
