

The ESX-5 System of Pathogenic Mycobacteria Is Involved In Capsule Integrity and Virulence through Its Substrate PPE10
- PMID: 27280885
- PMCID: PMC4900558
- DOI: 10.1371/journal.ppat.1005696
The ESX-5 System of Pathogenic Mycobacteria Is Involved In Capsule Integrity and Virulence through Its Substrate PPE10
Abstract
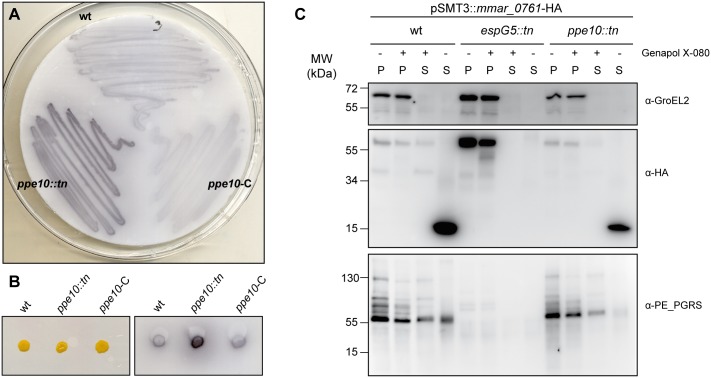
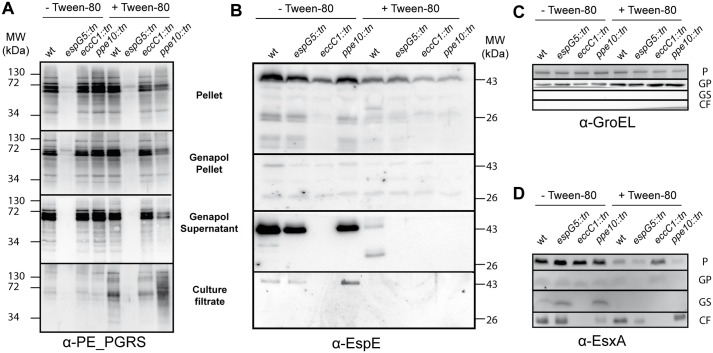
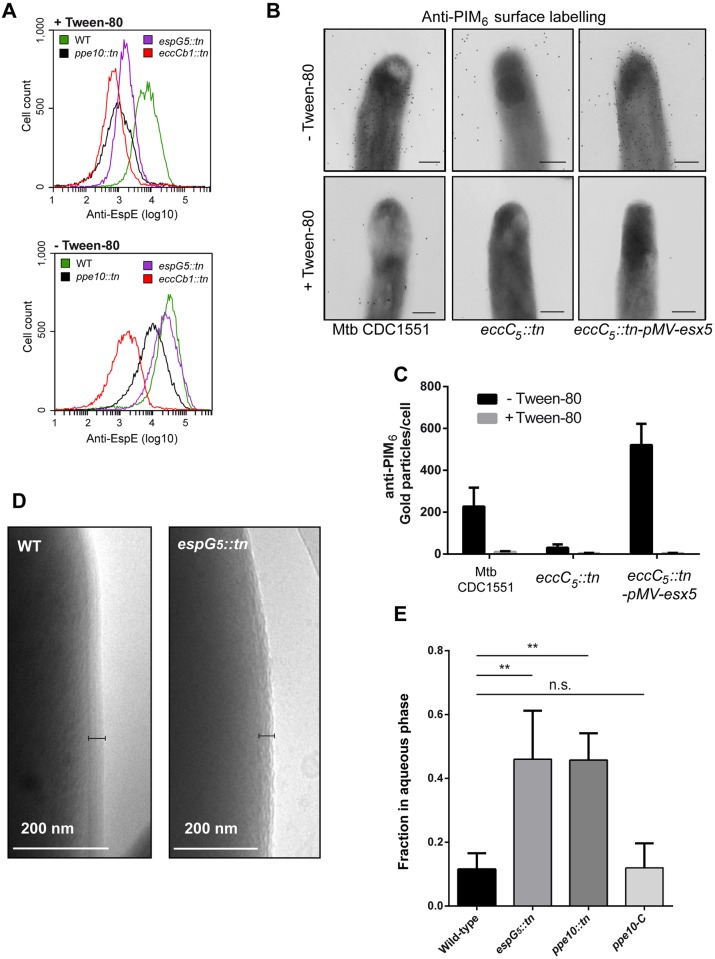
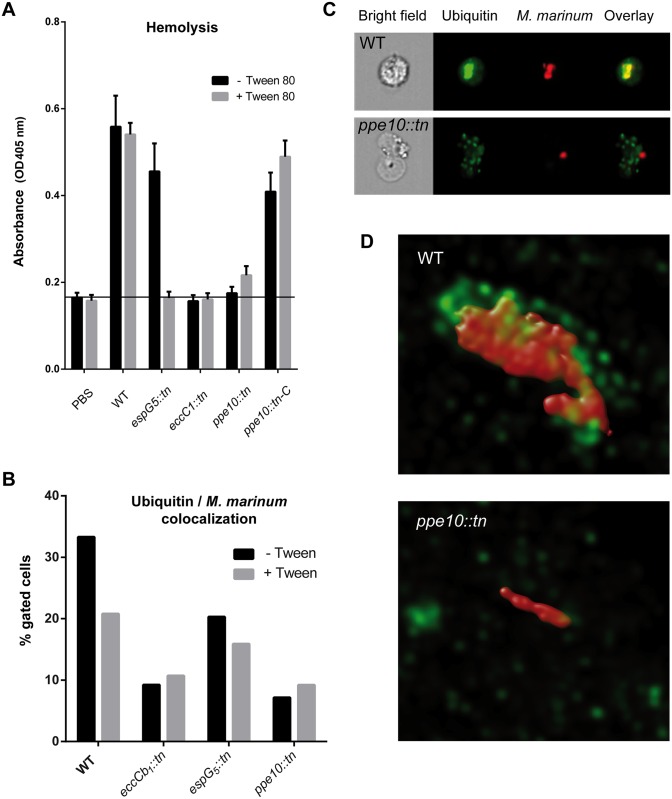
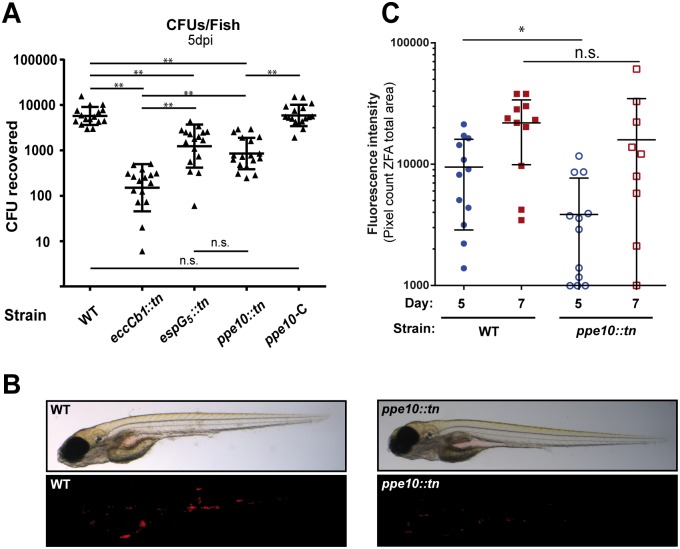
Mycobacteria produce a capsule layer, which consists of glycan-like polysaccharides and a number of specific proteins. In this study, we show that, in slow-growing mycobacteria, the type VII secretion system ESX-5 plays a major role in the integrity and stability of the capsule. We have identified PPE10 as the ESX-5 substrate responsible for this effect. Mutants in esx-5 and ppe10 both have impaired capsule integrity as well as reduced surface hydrophobicity. Electron microscopy, immunoblot and flow cytometry analyses demonstrated reduced amounts of surface localized proteins and glycolipids, and morphological differences in the capsular layer. Since capsular proteins secreted by the ESX-1 system are important virulence factors, we tested the effect of the mutations that cause capsular defects on virulence mechanisms. Both esx-5 and ppe10 mutants of Mycobacterium marinum were shown to be impaired in ESX-1-dependent hemolysis. In agreement with this, the ppe10 and esx5 mutants showed reduced recruitment of ubiquitin in early macrophage infection and intermediate attenuation in zebrafish embryos. These results provide a pivotal role for the ESX-5 secretion system and its substrate PPE10, in the capsular integrity of pathogenic mycobacteria. These findings open up new roads for research on the mycobacterial capsule and its role in virulence and immune modulation.
Conflict of interest statement
MM is employed by Eyen SE, but did not receive funding for his collaboration on this project. Eyen SE has not been financially involved in the development of this manuscript or the data and methods therein. These data do not financially benefit Eyen SE by any products, patents or other interests. This does not alter our adherence to all PLOS Pathogens policies on sharing data and materials. The other authors have declared that no competing interests exist.
Figures
References
-
- World Health Organization. Global tuberculosis report 2013. 2013.
-
- Brennan PJ, Crick DC. The cell-wall core of Mycobacterium tuberculosis in the context of drug discovery. Curr Top Med Chem. 2007;7: 475–88. Available: http://www.ncbi.nlm.nih.gov/pubmed/17346193 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
