

Stalled replication fork rescue requires a novel DNA helicase
- PMID: 27282357
- PMCID: PMC5040163
- DOI: 10.1016/j.ymeth.2016.06.002
Stalled replication fork rescue requires a novel DNA helicase
Abstract
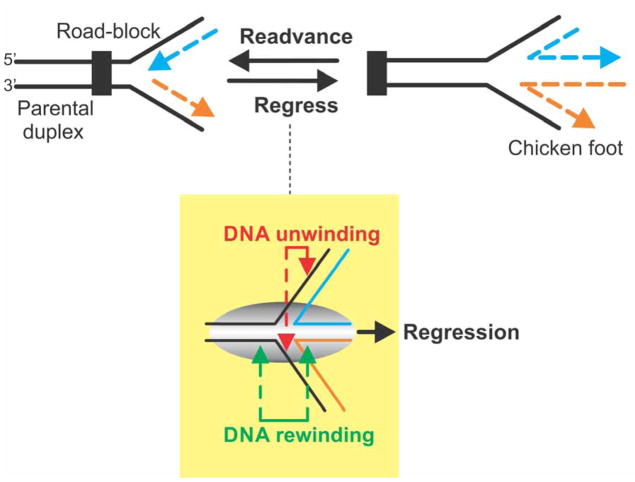
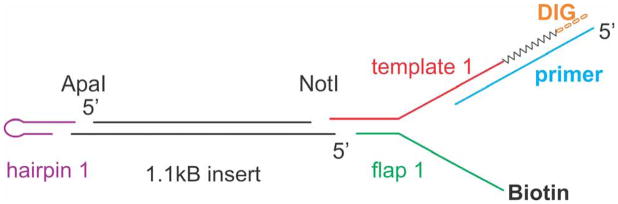
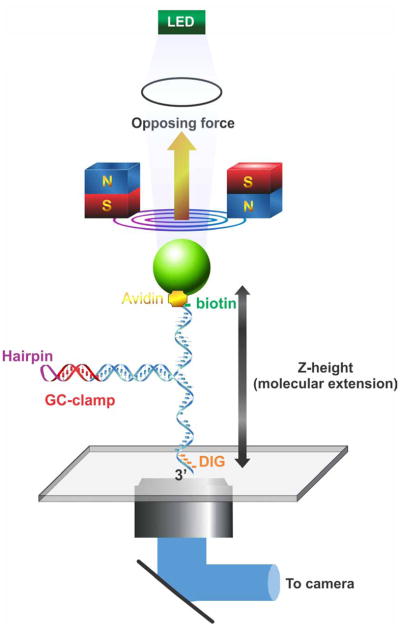
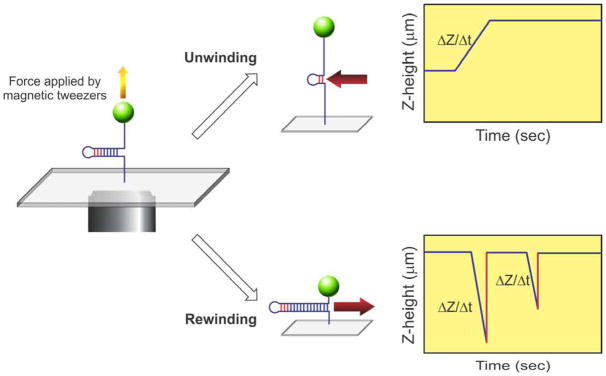
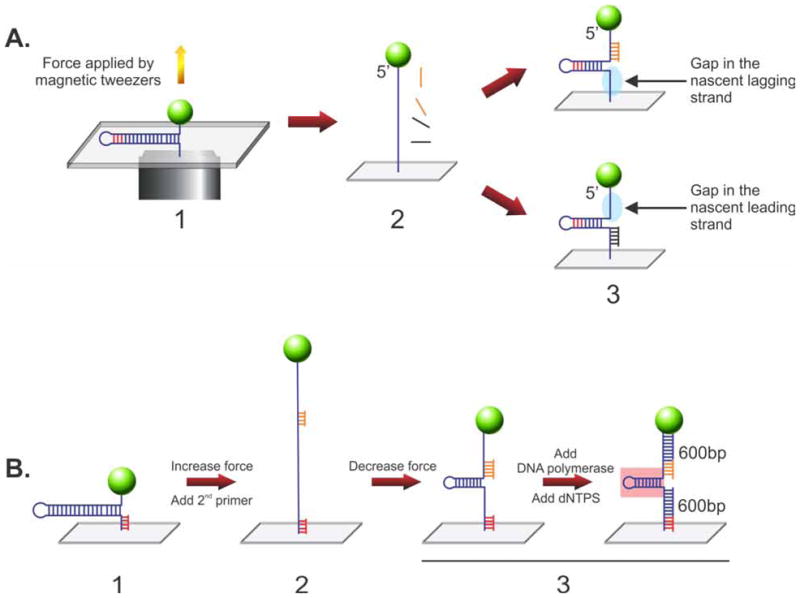
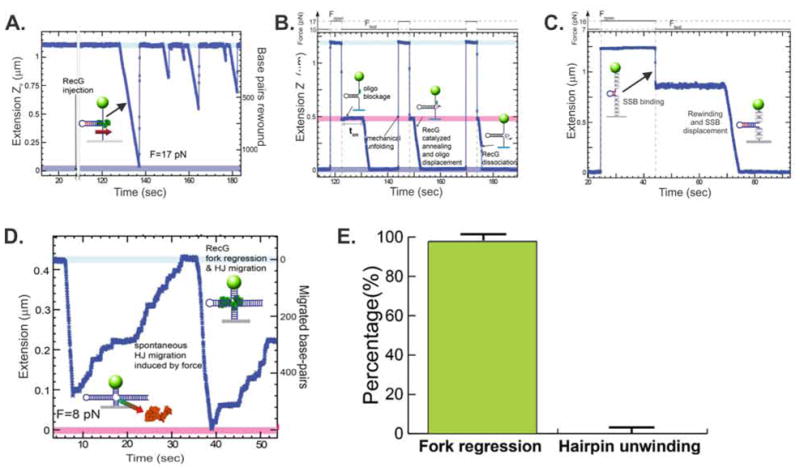
During DNA replication, forks often stall and require restart. One mechanism for restart requires that the fork be moved in a direction opposite to that of replication. This reaction is known as fork regression. For this reaction to occur, the enzyme must couple unwinding of the nascent heteroduplex fork arms to the rewinding of nascent strands ahead of itself and to the parental duplex in its wake. As the arms of the fork are complementary, this reaction is isoenergetic making it challenging to study. To overcome this, a novel adaptation of magnetic tweezers was developed by the Croquette group. Here, a 1200bp hairpin was attached at opposite ends to a flow cell surface and a magnetic bead. By manipulating the bead with the magnets, force can be applied to unwind the hairpin or alternatively, released to allow the hairpin to rewind. This adaptation was used to study fork regression by RecG. The results show that this is an efficient regression enzyme, able to work against a large opposing force. Critically, it couples DNA unwinding to duplex rewinding and in the process, can displace bound proteins from fork arms.
Keywords: DNA helicase; Fork regression; Hairpin substrate; Holliday junction; Magnetic tweezers; RecG; Replication fork; SSB.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
DNA Helicase-SSB Interactions Critical to the Regression and Restart of Stalled DNA Replication forks in Escherichia coli.Genes (Basel). 2020 Apr 26;11(5):471. doi: 10.3390/genes11050471. Genes (Basel). 2020. PMID: 32357475 Free PMC article. Review.
-
Rescue of stalled replication forks by RecG: simultaneous translocation on the leading and lagging strand templates supports an active DNA unwinding model of fork reversal and Holliday junction formation.Proc Natl Acad Sci U S A. 2001 Jul 17;98(15):8227-34. doi: 10.1073/pnas.111008698. Proc Natl Acad Sci U S A. 2001. PMID: 11459957 Free PMC article.
-
Remodeling of RecG Helicase at the DNA Replication Fork by SSB Protein.Sci Rep. 2015 Apr 29;5:9625. doi: 10.1038/srep09625. Sci Rep. 2015. PMID: 25923319 Free PMC article.
-
Escherichia coli PriA helicase: fork binding orients the helicase to unwind the lagging strand side of arrested replication forks.J Mol Biol. 2001 Oct 5;312(5):935-47. doi: 10.1006/jmbi.2001.4930. J Mol Biol. 2001. PMID: 11580240
-
The Biochemical Mechanism of Fork Regression in Prokaryotes and Eukaryotes-A Single Molecule Comparison.Int J Mol Sci. 2022 Aug 3;23(15):8613. doi: 10.3390/ijms23158613. Int J Mol Sci. 2022. PMID: 35955746 Free PMC article. Review.
Cited by
-
DNA Helicase-SSB Interactions Critical to the Regression and Restart of Stalled DNA Replication forks in Escherichia coli.Genes (Basel). 2020 Apr 26;11(5):471. doi: 10.3390/genes11050471. Genes (Basel). 2020. PMID: 32357475 Free PMC article. Review.
-
Rep and UvrD Antagonize One Another at Stalled Replication Forks and This Is Exacerbated by SSB.ACS Omega. 2019 Mar 31;4(3):5180-5196. doi: 10.1021/acsomega.8b02375. Epub 2019 Mar 12. ACS Omega. 2019. PMID: 30949615 Free PMC article.
-
SSB and the RecG DNA helicase: an intimate association to rescue a stalled replication fork.Protein Sci. 2017 Apr;26(4):638-649. doi: 10.1002/pro.3114. Epub 2017 Mar 17. Protein Sci. 2017. PMID: 28078722 Free PMC article. Review.
-
Special Methods collection on DNA helicases.Methods. 2016 Oct 1;108:1-3. doi: 10.1016/j.ymeth.2016.08.009. Epub 2016 Aug 24. Methods. 2016. PMID: 27565743 Free PMC article.
References
-
- Kowalczykowski SC. Initiation of genetic recombination and recombination-dependent replication. Trends in Biochemical Sciences. 2000;25(4):156–65. - PubMed
-
- Seigneur M, Bidnenko V, Ehrlich S, Michel B. RuvAB acts at arrested replication forks. Cell. 1998;95(3):419–30. - PubMed
-
- Cox MM, Goodman MF, Kreuzer KN, Sherratt DJ, Sandler SJ, Marians KJ. The importance of repairing stalled replication forks. Nature. 2000;404(6773):37–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
