

Curcumin inhibits HIV-1 by promoting Tat protein degradation
- PMID: 27283735
- PMCID: PMC4901322
- DOI: 10.1038/srep27539
Curcumin inhibits HIV-1 by promoting Tat protein degradation
Abstract
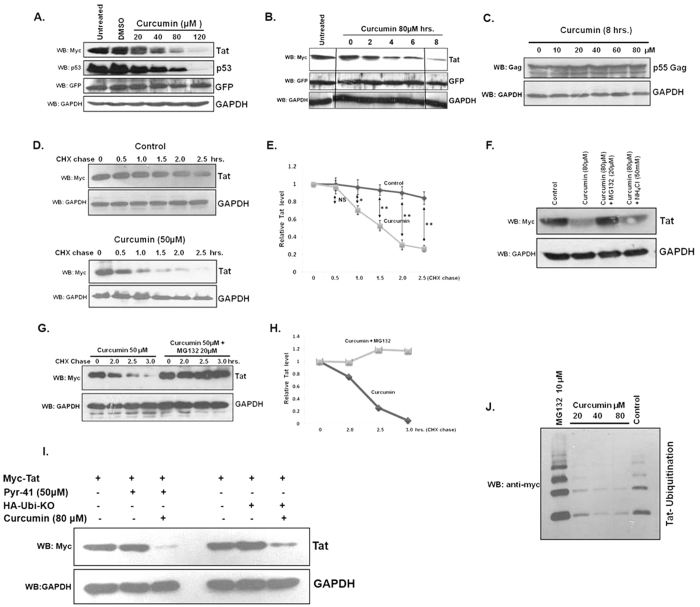
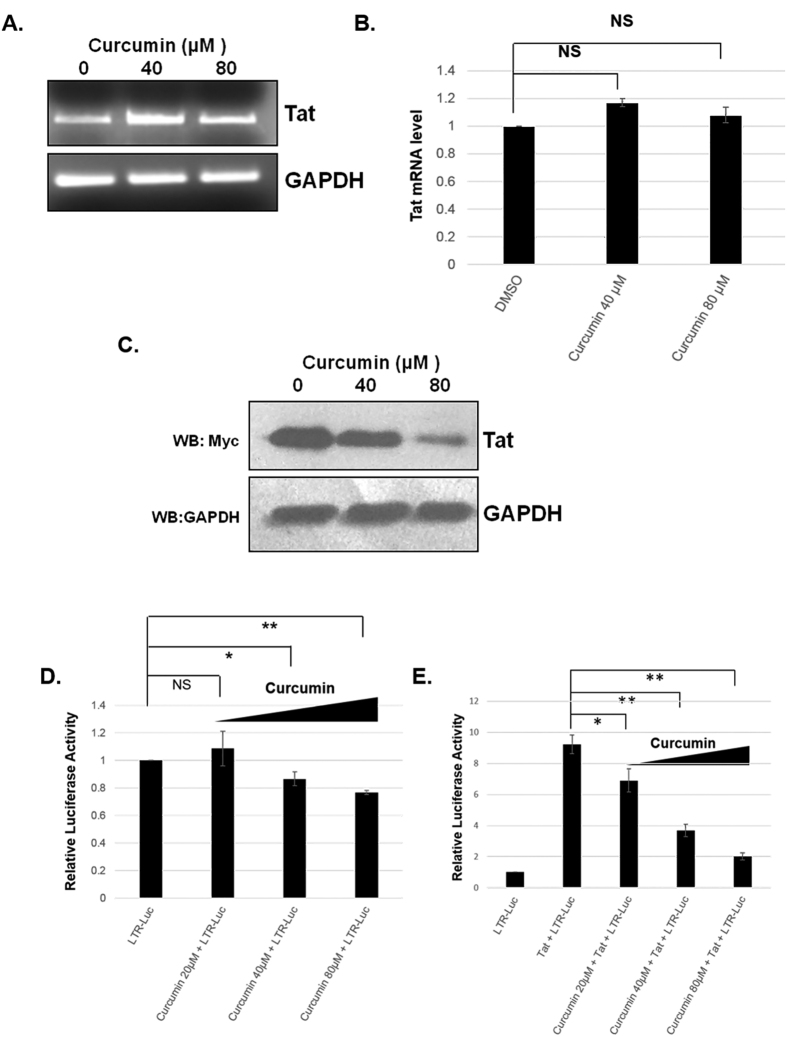
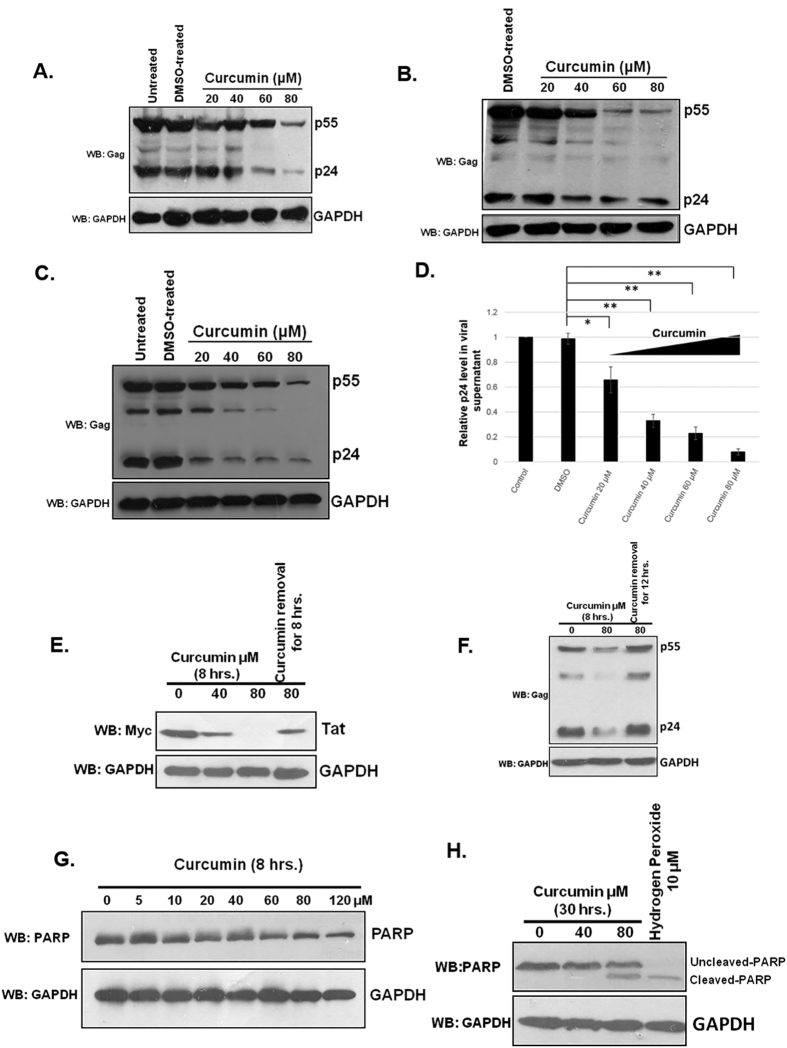
HIV-1 Tat is an intrinsically unfolded protein playing a pivotal role in viral replication by associating with TAR region of viral LTR. Unfolded proteins are degraded by 20S proteasome in an ubiquitin independent manner. Curcumin is known to activate 20S proteasome and promotes the degradation of intrinsically unfolded p53 tumor suppressor protein. Since HIV-1 Tat protein is largerly unfolded, we hypothesized that Tat may also be targeted through this pathway. Curcumin treated Tat transfected HEK-293T cells showed a dose and time dependent degradation of Tat protein. Contrary to this HIV-1 Gag which is a properly folded protein, remained unaffected with curcumin. Semi-quantitative RT-PCR analysis showed that curcumin treatment did not affect Tat gene transcription. Curcumin increased the rate of Tat protein degradation as shown by cycloheximide (CHX) chase assay. Degradation of the Tat protein is accomplished through proteasomal pathway as proteasomal inhibitor MG132 blocked Tat degradation. Curcumin also decreased Tat mediated LTR promoter transactivation and inhibited virus production from HIV-1 infected cells. Taken together our study reveals a novel observation that curcumin causes potent degradation of Tat which may be one of the major mechanisms behind its anti HIV activity.
Figures
References
-
- Ciechanover A. Intracellular protein degradation: from a vague idea thru the lysosome and the ubiquitin-proteasome system and onto human diseases and drug targeting. Cell Death Differ. 12, 1178–1190 (2005). - PubMed
-
- Hochstrasser M. Ubiquitin-dependent protein degradation. Annu. Rev. Genet. 30, 405–439 (1996). - PubMed
-
- Voges D., Zwickl P. & Baumeister W. The 26S proteasome: a molecular machine designed for controlled proteolysis. Annu. Rev. Biochem. 68, 1015–1068 (1999). - PubMed
-
- Asher G., Reuven N. & Shaul Y. 20S proteasomes and protein degradation “by default”. Bioessays 28, 844–849 (2006). - PubMed
-
- Jariel-Encontre I., Bossis G. & Piechaczyk M. Ubiquitin-independent degradation of proteins by the proteasome. Biochim. Biophys. Acta 1786, 153–177 (2008). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
