

Control of diverse subcellular processes by a single multi-functional lipid phosphatidylinositol 4,5-bisphosphate [PI(4,5)P2]
- PMID: 27288030
- PMCID: PMC6609453
- DOI: 10.1042/BCJ20160069
Control of diverse subcellular processes by a single multi-functional lipid phosphatidylinositol 4,5-bisphosphate [PI(4,5)P2]
Abstract
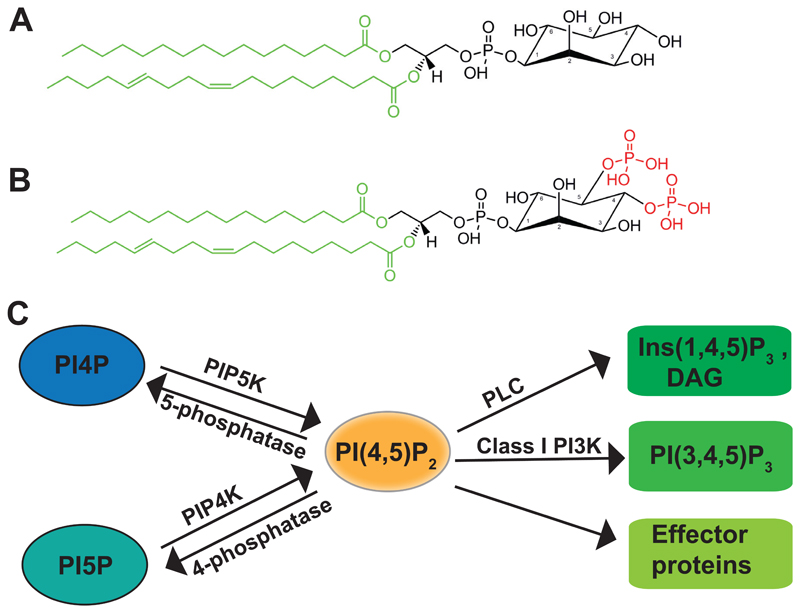
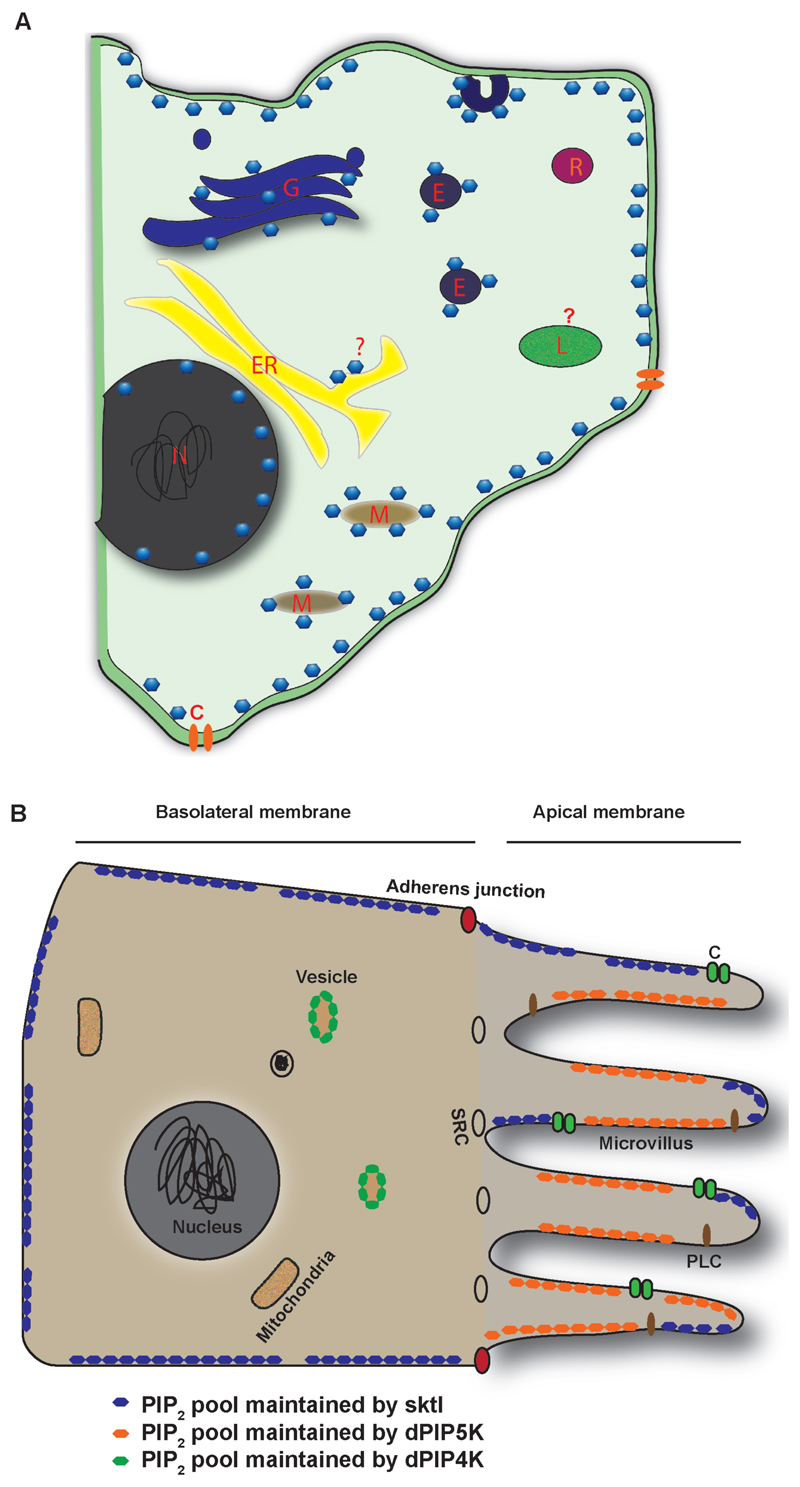
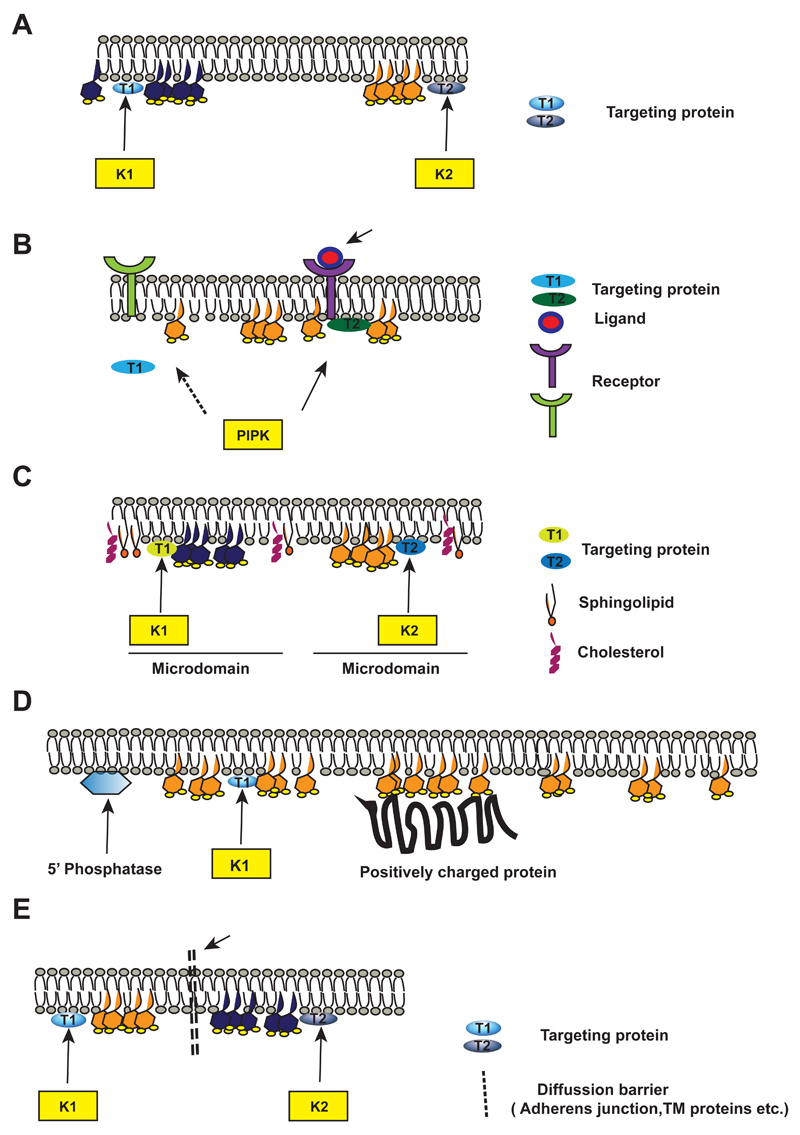
Phosphatidylinositol 4,5-bisphosphate [PI(4,5)P2] is a multi-functional lipid that regulates several essential subcellular processes in eukaryotic cells. In addition to its well-established function as a substrate for receptor-activated signalling at the plasma membrane (PM), it is now recognized that distinct PI(4,5)P2 pools are present at other organelle membranes. However, a long-standing question that remains unresolved is the mechanism by which a single lipid species, with an invariant functional head group, delivers numerous functions without loss of fidelity. In the present review, we summarize studies that have examined the molecular processes that shape the repertoire of PI(4,5)P2 pools in diverse eukaryotes. Collectively, these studies indicate a conserved role for lipid kinase isoforms in generating functionally distinct pools of PI(4,5)P2 in diverse metazoan species. The sophistication underlying the regulation of multiple functions by PI(4,5)P2 is also shaped by mechanisms that regulate its availability to enzymes involved in its metabolism as well as molecular processes that control its diffusion at nanoscales in the PM. Collectively, these mechanisms ensure the specificity of PI(4,5)P2 mediated signalling at eukaryotic membranes.
Keywords: 5)P2 pools; PI(4; cell membranes; lipid kinase; lipid microdomains; phosphoinositides; phospholipase C (PLC) signalling.
© 2016 The Author(s). published by Portland Press Limited on behalf of the Biochemical Society.
Figures
References
-
- Pfeffer S, Aivazian D. Targeting Rab GTPases to distinct membrane compartments. Nat Rev Mol Cell Biol. 2004;5:886–96. - PubMed
-
- Jost M, Simpson F, Kavran JM, Lemmon Ma, Schmid SL. Phosphatidylinositol-4,5-bisphosphate is required for endocytic coated vesicle formation. Curr Biol. 8:1399–1402. - PubMed
-
- Ford MG, Pearse BM, Higgins MK, Vallis Y, Owen DJ, Gibson a, Hopkins CR, Evans PR, McMahon HT. Simultaneous binding of PtdIns(4,5)P2 and clathrin by AP180 in the nucleation of clathrin lattices on membranes. Science. 2001;291:1051–1055. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous