

Structural basis for the antagonistic roles of RNP-8 and GLD-3 in GLD-2 poly(A)-polymerase activity
- PMID: 27288313
- PMCID: PMC4931106
- DOI: 10.1261/rna.056598.116
Structural basis for the antagonistic roles of RNP-8 and GLD-3 in GLD-2 poly(A)-polymerase activity
Abstract
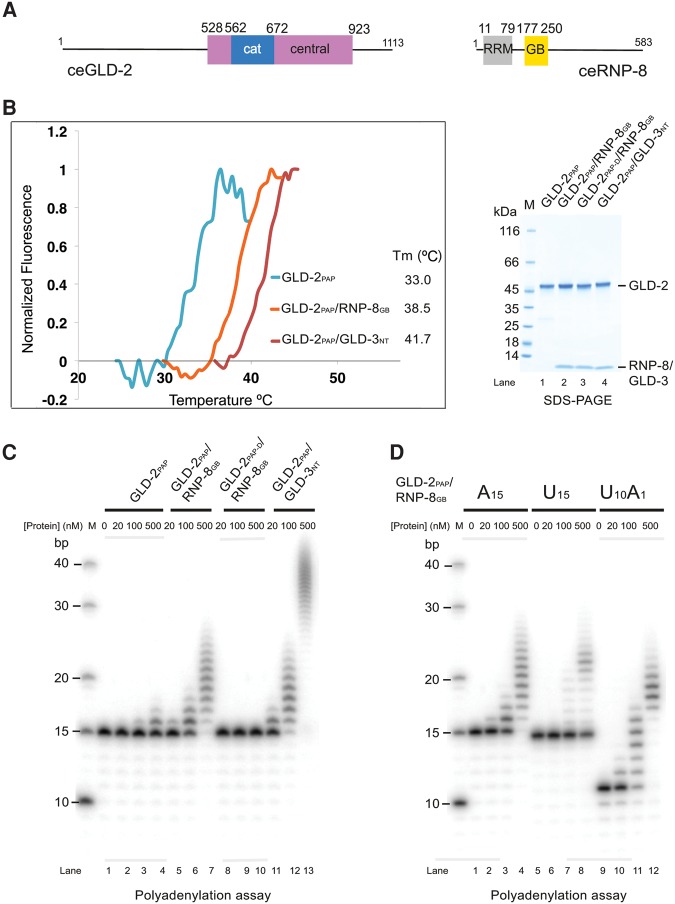
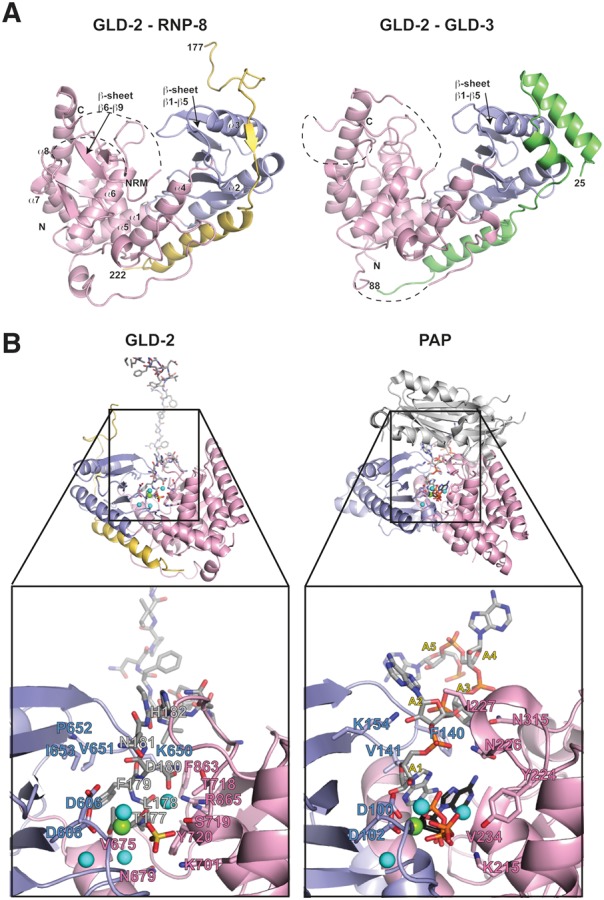
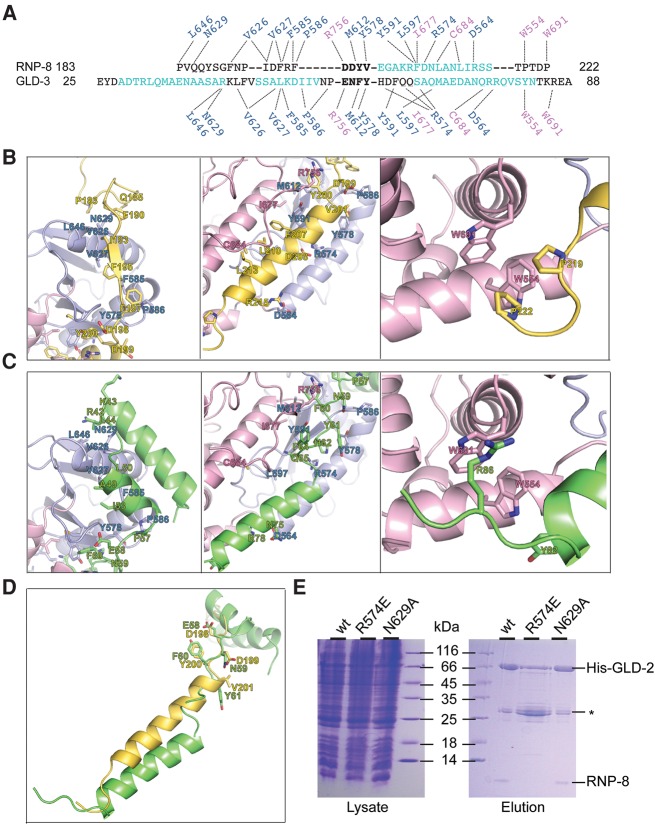
Cytoplasmic polyadenylation drives the translational activation of specific mRNAs in early metazoan development and is performed by distinct complexes that share the same catalytic poly(A)-polymerase subunit, GLD-2. The activity and specificity of GLD-2 depend on its binding partners. In Caenorhabditis elegans, GLD-2 promotes spermatogenesis when bound to GLD-3 and oogenesis when bound to RNP-8. GLD-3 and RNP-8 antagonize each other and compete for GLD-2 binding. Following up on our previous mechanistic studies of GLD-2-GLD-3, we report here the 2.5 Å resolution structure and biochemical characterization of a GLD-2-RNP-8 core complex. In the structure, RNP-8 embraces the poly(A)-polymerase, docking onto several conserved hydrophobic hotspots present on the GLD-2 surface. RNP-8 stabilizes GLD-2 and indirectly stimulates polyadenylation. RNP-8 has a different amino-acid sequence and structure as compared to GLD-3. Yet, it binds the same surfaces of GLD-2 by forming alternative interactions, rationalizing the remarkable versatility of GLD-2 complexes.
Keywords: C. elegans; cytoplasmic polyadenylation; germline development; nucleotidyl-transferase; translational regulation.
© 2016 Nakel et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
Similar articles
-
Structural basis for the activation of the C. elegans noncanonical cytoplasmic poly(A)-polymerase GLD-2 by GLD-3.Proc Natl Acad Sci U S A. 2015 Jul 14;112(28):8614-9. doi: 10.1073/pnas.1504648112. Epub 2015 Jun 29. Proc Natl Acad Sci U S A. 2015. PMID: 26124149 Free PMC article.
-
Antagonism between GLD-2 binding partners controls gamete sex.Dev Cell. 2009 May;16(5):723-33. doi: 10.1016/j.devcel.2009.04.002. Dev Cell. 2009. PMID: 19460348 Free PMC article.
-
A regulatory cytoplasmic poly(A) polymerase in Caenorhabditis elegans.Nature. 2002 Sep 19;419(6904):312-6. doi: 10.1038/nature01039. Nature. 2002. PMID: 12239571
-
Poly(A) tail synthesis and regulation: recent structural insights.Curr Opin Struct Biol. 2002 Feb;12(1):82-8. doi: 10.1016/s0959-440x(02)00293-2. Curr Opin Struct Biol. 2002. PMID: 11839494 Review.
-
The regulatory network controlling the proliferation-meiotic entry decision in the Caenorhabditis elegans germ line.Curr Top Dev Biol. 2006;76:185-215. doi: 10.1016/S0070-2153(06)76006-9. Curr Top Dev Biol. 2006. PMID: 17118267 Review.
Cited by
-
Functions and mechanisms of RNA tailing by metazoan terminal nucleotidyltransferases.Wiley Interdiscip Rev RNA. 2021 Mar;12(2):e1622. doi: 10.1002/wrna.1622. Epub 2020 Jul 22. Wiley Interdiscip Rev RNA. 2021. PMID: 33145994 Free PMC article. Review.
-
LIN-41 and OMA Ribonucleoprotein Complexes Mediate a Translational Repression-to-Activation Switch Controlling Oocyte Meiotic Maturation and the Oocyte-to-Embryo Transition in Caenorhabditis elegans.Genetics. 2017 Aug;206(4):2007-2039. doi: 10.1534/genetics.117.203174. Epub 2017 Jun 1. Genetics. 2017. PMID: 28576864 Free PMC article.
-
Gld2 activity and RNA specificity is dynamically regulated by phosphorylation and interaction with QKI-7.RNA Biol. 2021 Oct 15;18(sup1):397-408. doi: 10.1080/15476286.2021.1952540. Epub 2021 Jul 21. RNA Biol. 2021. PMID: 34288801 Free PMC article.
-
Germline immortality relies on TRIM32-mediated turnover of a maternal mRNA activator in C. elegans.Sci Adv. 2022 Oct 14;8(41):eabn0897. doi: 10.1126/sciadv.abn0897. Epub 2022 Oct 14. Sci Adv. 2022. PMID: 36240265 Free PMC article.
-
Gld2 activity is regulated by phosphorylation in the N-terminal domain.RNA Biol. 2019 Aug;16(8):1022-1033. doi: 10.1080/15476286.2019.1608754. Epub 2019 May 5. RNA Biol. 2019. PMID: 31057087 Free PMC article.
References
-
- Barnard DC, Ryan K, Manley JL, Richter JD. 2004. Symplekin and xGLD-2 are required for CPEB-mediated cytoplasmic polyadenylation. Cell 119: 641–651. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources