

Cross-membranes orchestrate compartmentalization and morphogenesis in Streptomyces
- PMID: 27291257
- PMCID: PMC4909990
- DOI: 10.1038/ncomms11836
Cross-membranes orchestrate compartmentalization and morphogenesis in Streptomyces
Abstract
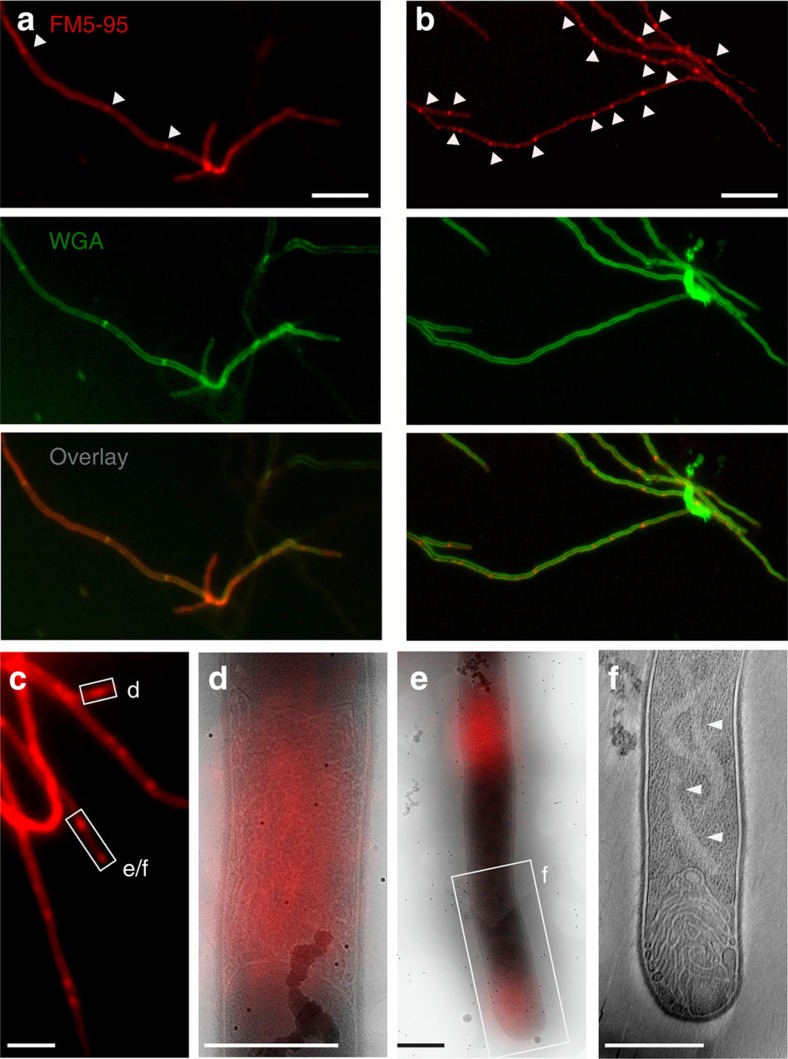
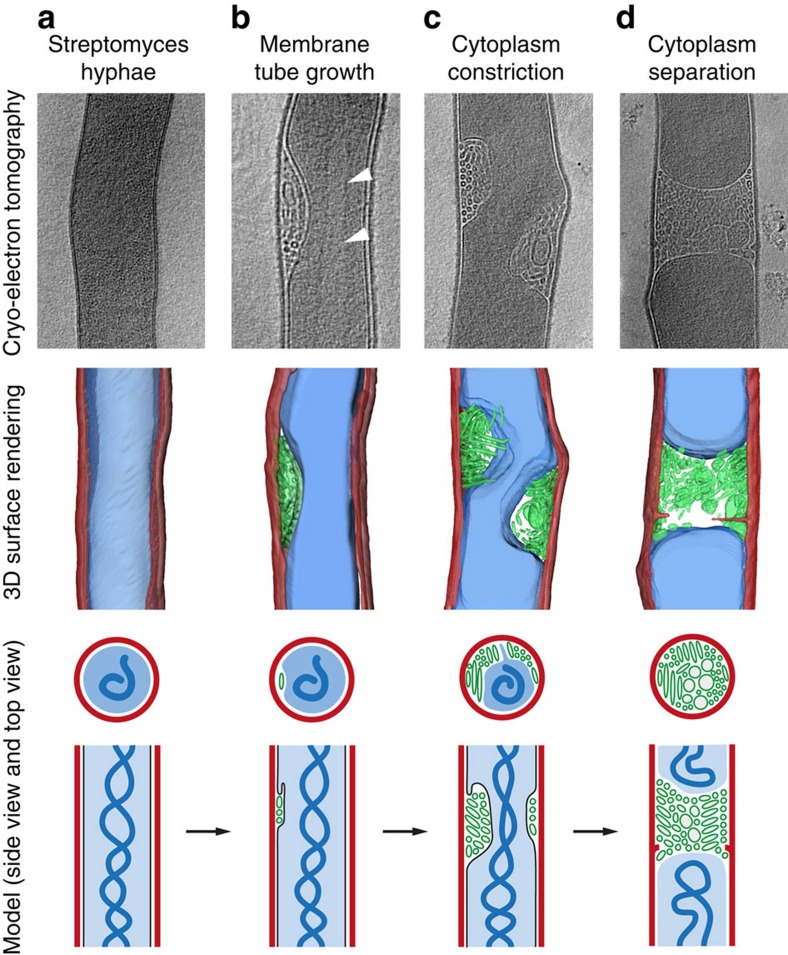
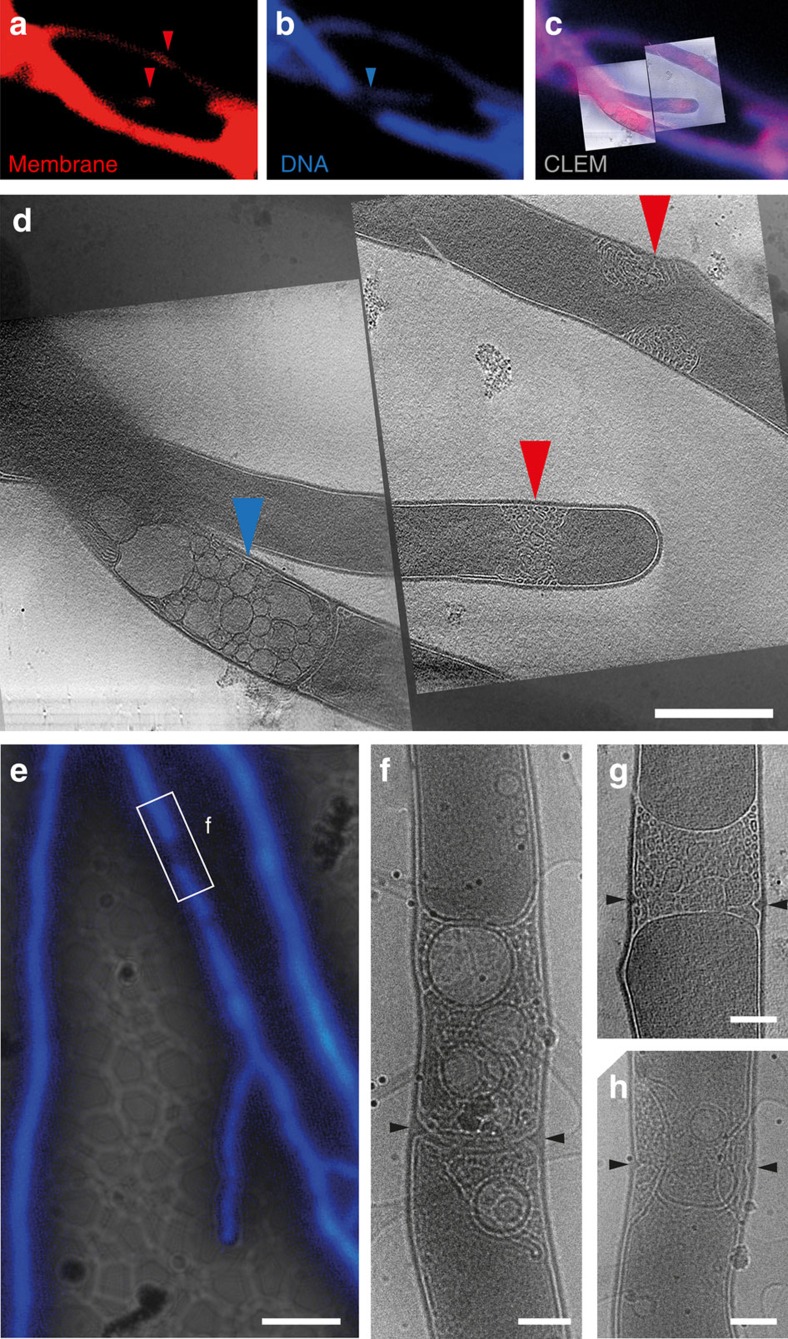
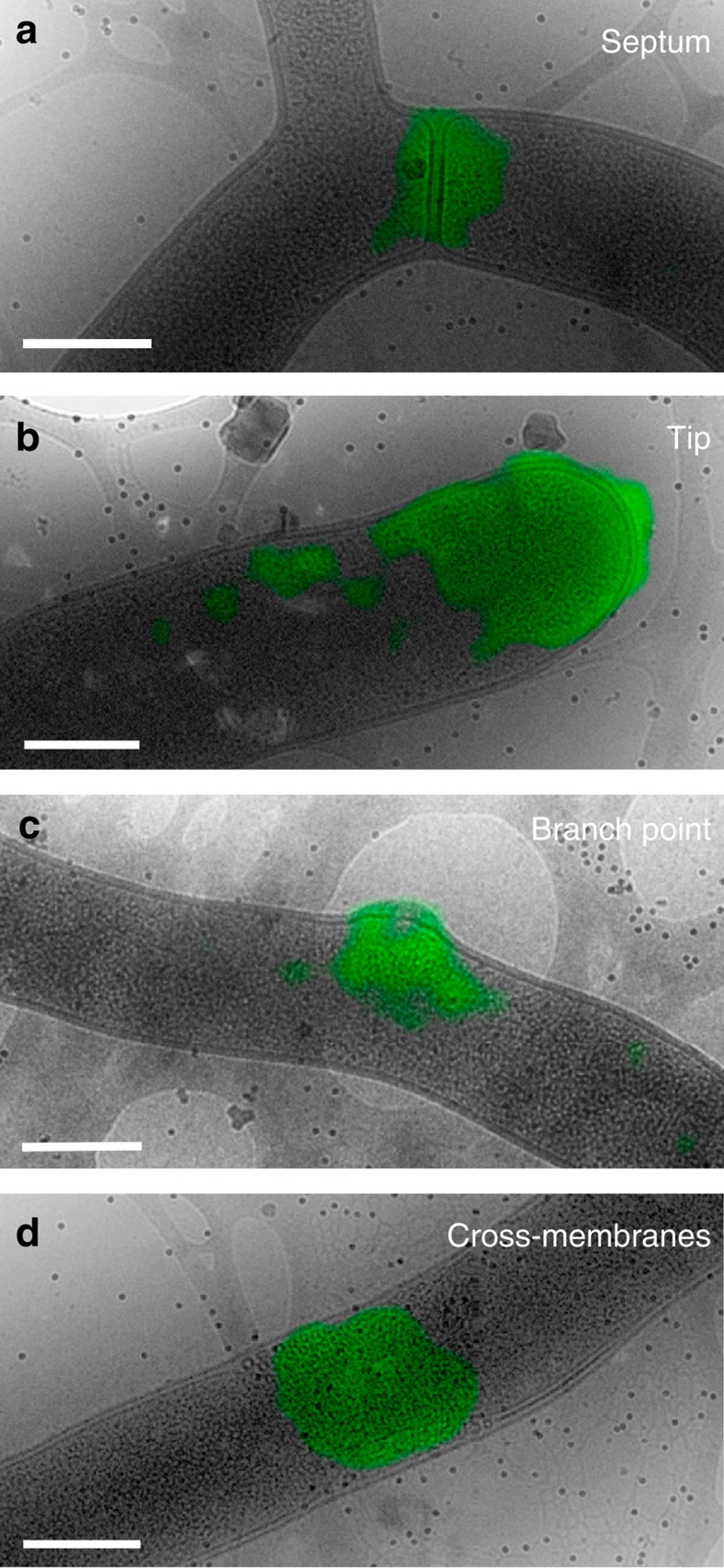
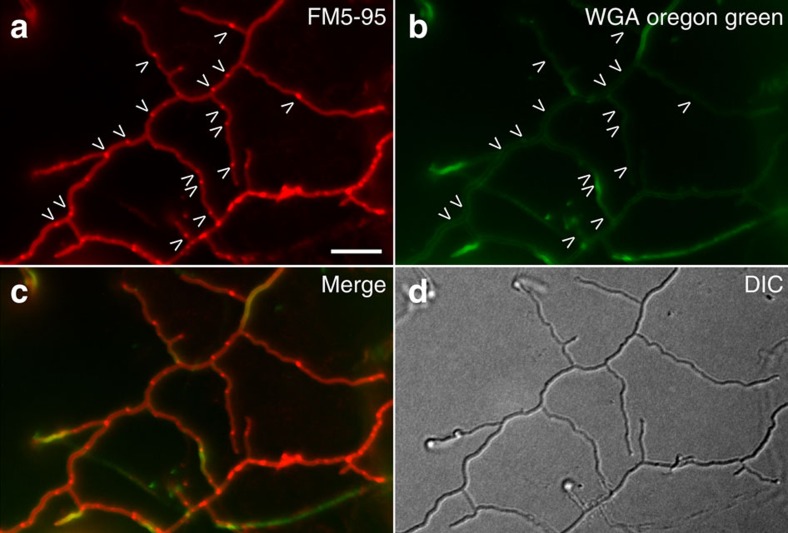
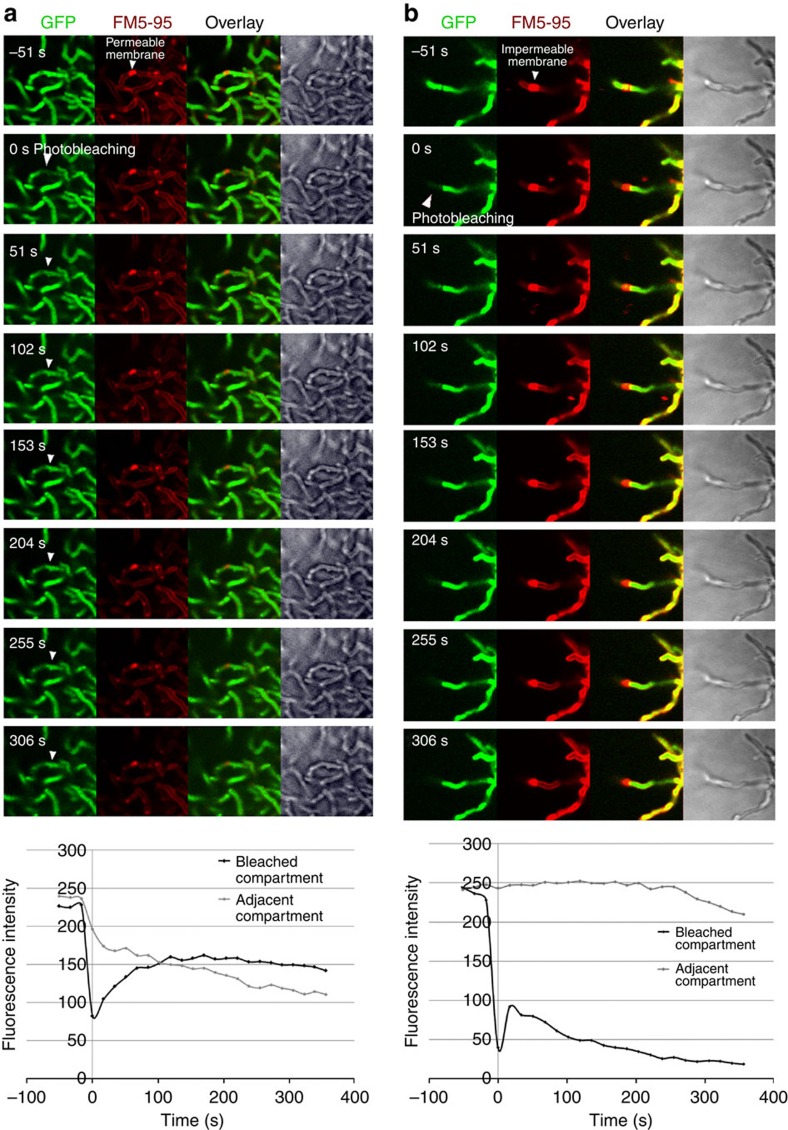
Far from being simple unicellular entities, bacteria have complex social behaviour and organization, living in large populations, and some even as coherent, multicellular entities. The filamentous streptomycetes epitomize such multicellularity, growing as a syncytial mycelium with physiologically distinct hyphal compartments separated by infrequent cross-walls. The viability of mutants devoid of cell division, which can be propagated from fragments, suggests the presence of a different form of compartmentalization in the mycelium. Here we show that complex membranes, visualized by cryo-correlative light microscopy and electron tomography, fulfil this role. Membranes form small assemblies between the cell wall and cytoplasmic membrane, or, as evidenced by FRAP experiments, large protein-impermeable cross-membrane structures, which compartmentalize the multinucleoid mycelium. All areas containing cross-membrane structures are nucleoid-restricted zones, suggesting that the membrane assemblies may also act to protect nucleoids from cell-wall restructuring events. Our work reveals a novel mechanism of controlling compartmentalization and development in multicellular bacteria.
Figures
References
-
- Shapiro J. A. Thinking about bacterial populations as multicellular organisms. Annu. Rev. Microbiol. 52, 81–104 (1998). - PubMed
-
- Claessen D., Rozen D. E., Kuipers O. P., Sogaard-Andersen L. & van Wezel G. P. Bacterial solutions to multicellularity: a tale of biofilms, filaments and fruiting bodies. Nat. Rev. Microbiol. 12, 115–124 (2014). - PubMed
-
- Miguelez E. M., Hardisson C. & Manzanal M. B. Streptomycetes: a new model to study cell death. Int. Microbiol. 3, 153–158 (2000). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
