

Partitioning the metabolic scope: the importance of anaerobic metabolism and implications for the oxygen- and capacity-limited thermal tolerance (OCLTT) hypothesis
- PMID: 27293766
- PMCID: PMC4896295
- DOI: 10.1093/conphys/cow019
Partitioning the metabolic scope: the importance of anaerobic metabolism and implications for the oxygen- and capacity-limited thermal tolerance (OCLTT) hypothesis
Abstract
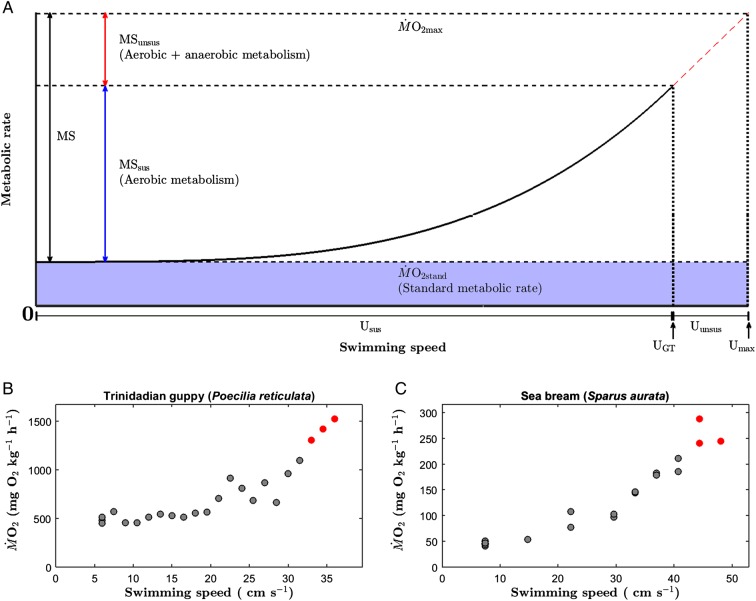
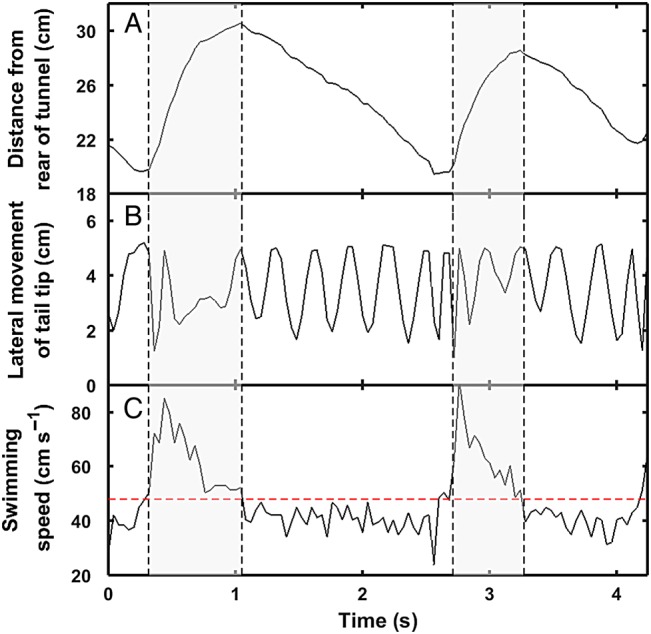
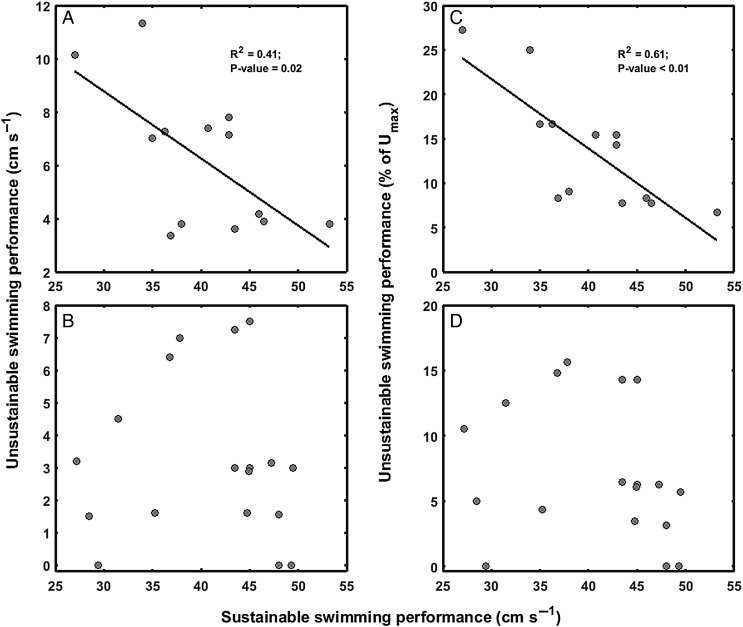
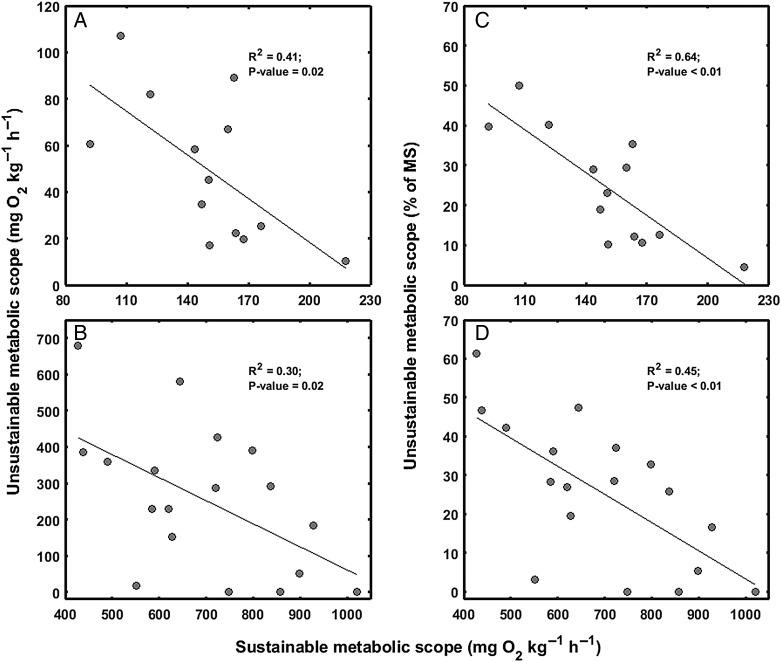
Ongoing climate change is predicted to affect the distribution and abundance of aquatic ectotherms owing to increasing constraints on organismal physiology, in particular involving the metabolic scope (MS) available for performance and fitness. The oxygen- and capacity-limited thermal tolerance (OCLTT) hypothesis prescribes MS as an overarching benchmark for fitness-related performance and assumes that any anaerobic contribution within the MS is insignificant. The MS is typically derived from respirometry by subtracting standard metabolic rate from the maximal metabolic rate; however, the methodology rarely accounts for anaerobic metabolism within the MS. Using gilthead sea bream (Sparus aurata) and Trinidadian guppy (Poecilia reticulata), this study tested for trade-offs (i) between aerobic and anaerobic components of locomotor performance; and (ii) between the corresponding components of the MS. Data collection involved measuring oxygen consumption rate at increasing swimming speeds, using the gait transition from steady to unsteady (burst-assisted) swimming to detect the onset of anaerobic metabolism. Results provided evidence of the locomotor performance trade-off, but only in S. aurata. In contrast, both species revealed significant negative correlations between aerobic and anaerobic components of the MS, indicating a trade-off where both components of the MS cannot be optimized simultaneously. Importantly, the fraction of the MS influenced by anaerobic metabolism was on average 24.3 and 26.1% in S. aurata and P. reticulata, respectively. These data highlight the importance of taking anaerobic metabolism into account when assessing effects of environmental variation on the MS, because the fraction where anaerobic metabolism occurs is a poor indicator of sustainable aerobic performance. Our results suggest that without accounting for anaerobic metabolism within the MS, studies involving the OCLTT hypothesis could overestimate the metabolic scope available for sustainable activities and the ability of individuals and species to cope with climate change.
Keywords: Aerobic metabolic scope; Trinidadian guppy (Poecilia reticulata); anaerobic metabolism; oxygen- and capacity-limited thermal tolerance (OCLTT); sea bream (Sparus aurata); trade-off.
Figures
References
-
- Alexander RM. (1989) Optimization and gaits in the locomotion of vertebrates. Physiol Rev 69: 1199–1227. - PubMed
-
- Altimiras J, Claireaux G, Sandblom E, Farrell AP, McKenzie DJ, Axelsson M (2008) Gastrointestinal blood flow and postprandial metabolism in swimming sea bass Dicentrarchus labrax. Physiol Biochem Zool 81: 663–672. - PubMed
-
- Anttila K, Jørgensen SM, Casselman MT, Timmerhaus G, Farrell AP, Takle H (2014) Association between swimming performance, cardiorespiratory morphometry, and thermal tolerance in Atlantic salmon (Salmo salar L.). Front Mar Sci 1: 76.
-
- Arnott SA, Chiba S, Conover DO (2006) Evolution of intrinsic growth rate: metabolic costs drive trade-offs between growth and swimming performance in Menidia menidia. Evolution 60: 1269–1278. - PubMed
-
- Binning SA, Roche DG, Fulton CJ (2014) Localised intraspecific variation in the swimming phenotype of a coral reef fish across different wave exposures. Oecologia 174: 623–630. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
