

Neural Representation of Concurrent Vowels in Macaque Primary Auditory Cortex
- PMID: 27294198
- PMCID: PMC4901243
- DOI: 10.1523/ENEURO.0071-16.2016
Neural Representation of Concurrent Vowels in Macaque Primary Auditory Cortex
Abstract
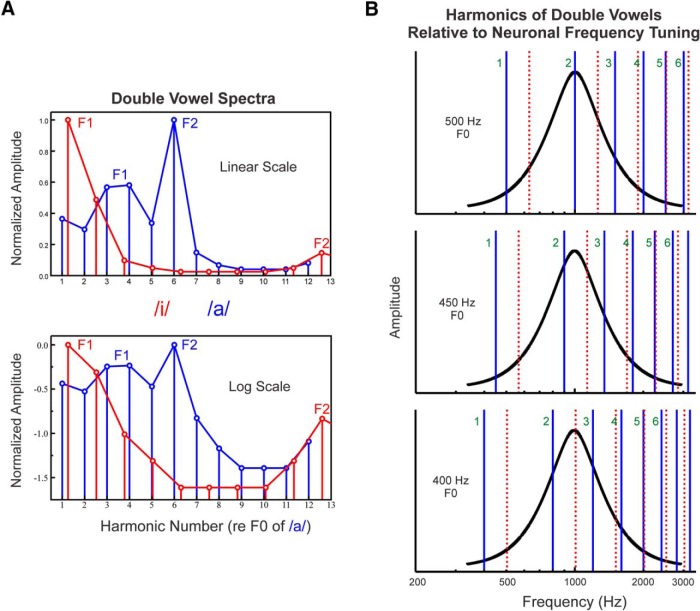
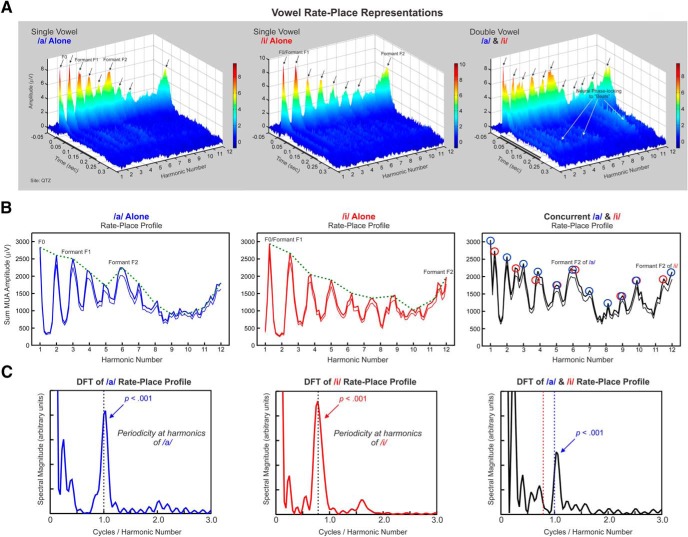
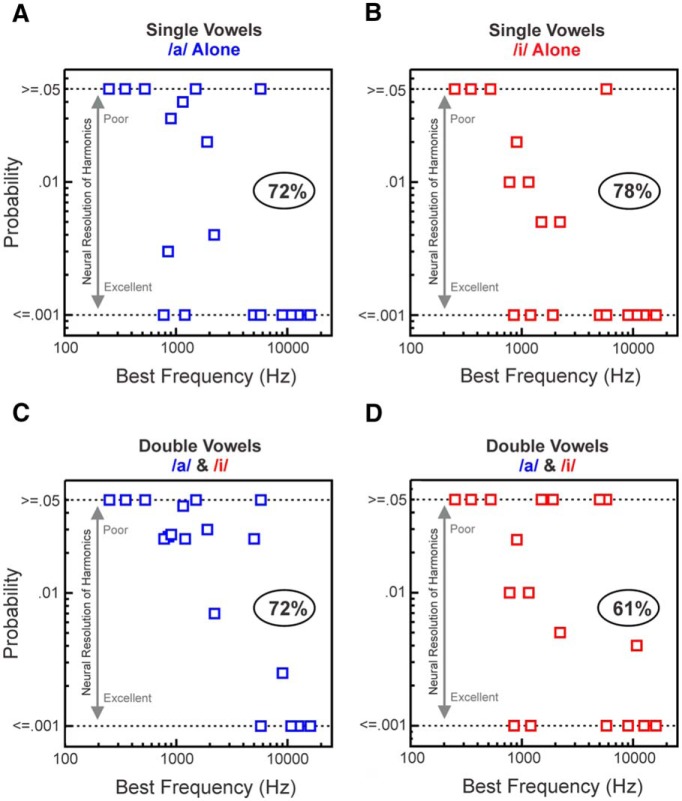
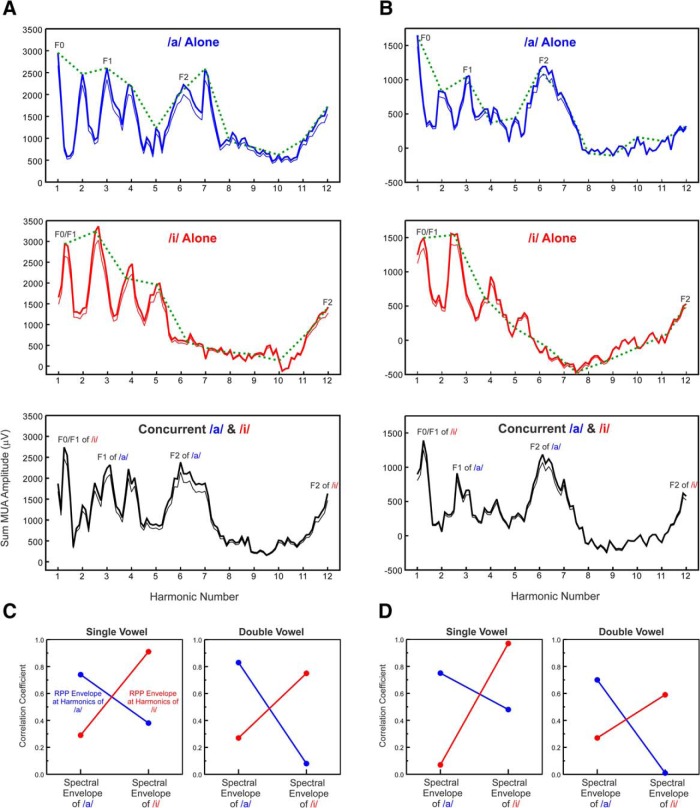
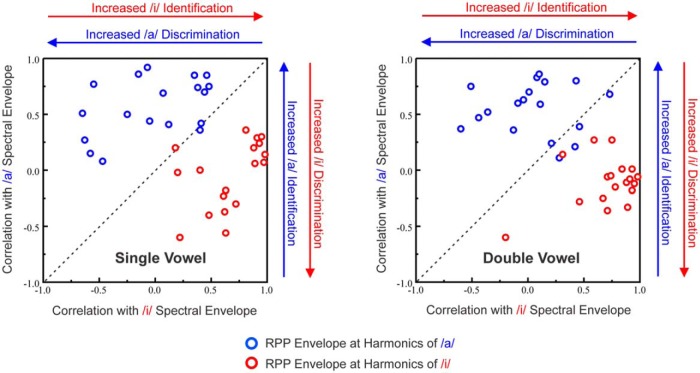
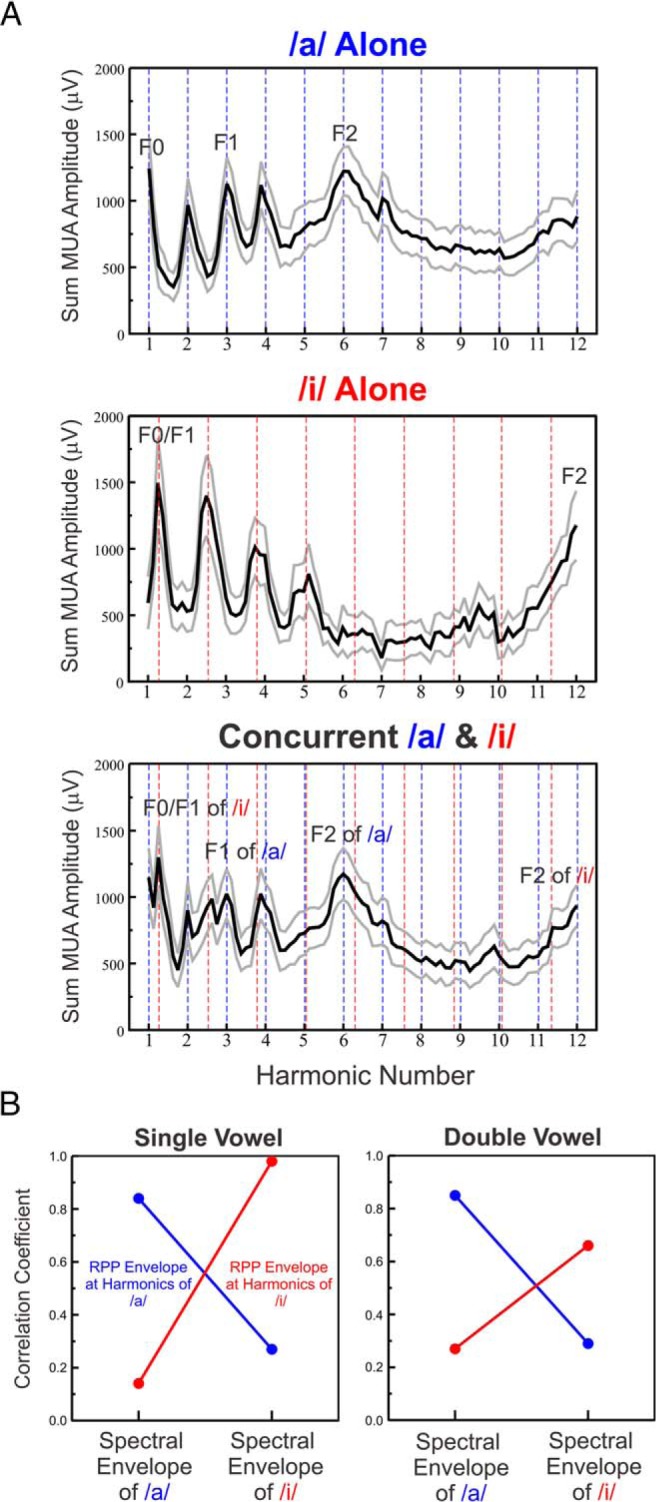
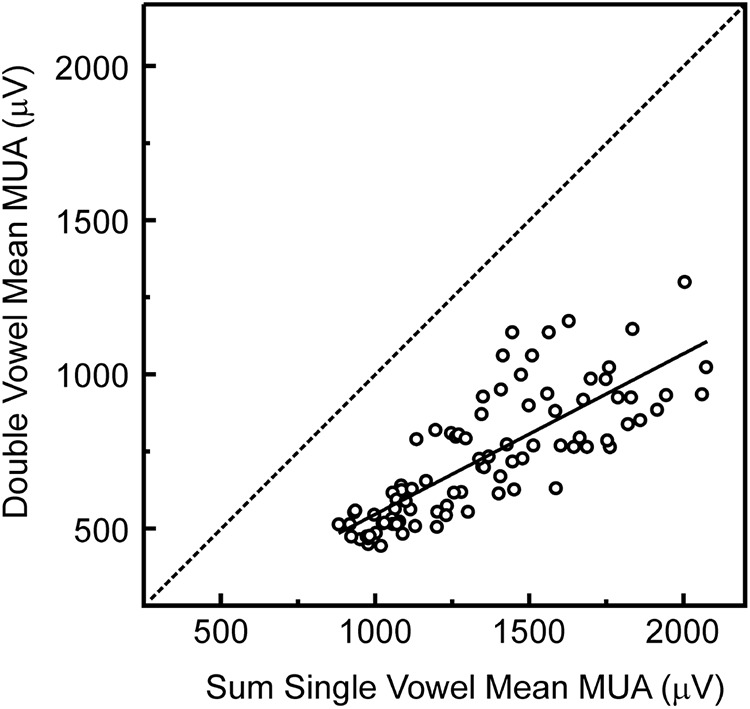
Successful speech perception in real-world environments requires that the auditory system segregate competing voices that overlap in frequency and time into separate streams. Vowels are major constituents of speech and are comprised of frequencies (harmonics) that are integer multiples of a common fundamental frequency (F0). The pitch and identity of a vowel are determined by its F0 and spectral envelope (formant structure), respectively. When two spectrally overlapping vowels differing in F0 are presented concurrently, they can be readily perceived as two separate "auditory objects" with pitches at their respective F0s. A difference in pitch between two simultaneous vowels provides a powerful cue for their segregation, which in turn, facilitates their individual identification. The neural mechanisms underlying the segregation of concurrent vowels based on pitch differences are poorly understood. Here, we examine neural population responses in macaque primary auditory cortex (A1) to single and double concurrent vowels (/a/ and /i/) that differ in F0 such that they are heard as two separate auditory objects with distinct pitches. We find that neural population responses in A1 can resolve, via a rate-place code, lower harmonics of both single and double concurrent vowels. Furthermore, we show that the formant structures, and hence the identities, of single vowels can be reliably recovered from the neural representation of double concurrent vowels. We conclude that A1 contains sufficient spectral information to enable concurrent vowel segregation and identification by downstream cortical areas.
Keywords: auditory scene analysis; multiunit activity; pitch; speech perception.
Conflict of interest statement
The authors report no conflict of interest.
Figures
Similar articles
-
Neural representation of concurrent harmonic sounds in monkey primary auditory cortex: implications for models of auditory scene analysis.J Neurosci. 2014 Sep 10;34(37):12425-43. doi: 10.1523/JNEUROSCI.0025-14.2014. J Neurosci. 2014. PMID: 25209282 Free PMC article.
-
Neural representation of harmonic complex tones in primary auditory cortex of the awake monkey.J Neurosci. 2013 Jun 19;33(25):10312-23. doi: 10.1523/JNEUROSCI.0020-13.2013. J Neurosci. 2013. PMID: 23785145 Free PMC article.
-
Noise and pitch interact during the cortical segregation of concurrent speech.Hear Res. 2017 Aug;351:34-44. doi: 10.1016/j.heares.2017.05.008. Epub 2017 May 25. Hear Res. 2017. PMID: 28578876
-
Neurophysiology and neuroanatomy of pitch perception: auditory cortex.Ann N Y Acad Sci. 2005 Dec;1060:148-74. doi: 10.1196/annals.1360.011. Ann N Y Acad Sci. 2005. PMID: 16597761 Review.
-
Pitch, harmonicity and concurrent sound segregation: psychoacoustical and neurophysiological findings.Hear Res. 2010 Jul;266(1-2):36-51. doi: 10.1016/j.heares.2009.09.012. Epub 2009 Sep 27. Hear Res. 2010. PMID: 19788920 Free PMC article. Review.
Cited by
-
Cortical tracking of voice pitch in the presence of multiple speakers depends on selective attention.Front Neurosci. 2022 Aug 8;16:828546. doi: 10.3389/fnins.2022.828546. eCollection 2022. Front Neurosci. 2022. PMID: 36003957 Free PMC article.
-
Functional characterization of human Heschl's gyrus in response to natural speech.Neuroimage. 2021 Jul 15;235:118003. doi: 10.1016/j.neuroimage.2021.118003. Epub 2021 Mar 28. Neuroimage. 2021. PMID: 33789135 Free PMC article.
-
Recent advances in understanding the auditory cortex.F1000Res. 2018 Sep 26;7:F1000 Faculty Rev-1555. doi: 10.12688/f1000research.15580.1. eCollection 2018. F1000Res. 2018. PMID: 30345008 Free PMC article. Review.
-
Monkeys share the neurophysiological basis for encoding sound periodicities captured by the frequency-following response with humans.Sci Rep. 2017 Nov 30;7(1):16687. doi: 10.1038/s41598-017-16774-8. Sci Rep. 2017. PMID: 29192170 Free PMC article.
-
Contribution of spiking activity in the primary auditory cortex to detection in noise.J Neurophysiol. 2017 Dec 1;118(6):3118-3131. doi: 10.1152/jn.00521.2017. Epub 2017 Aug 30. J Neurophysiol. 2017. PMID: 28855294 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources