

Genetic and biochemical evidence that gastrulation defects in Pofut2 mutants result from defects in ADAMTS9 secretion
- PMID: 27297885
- PMCID: PMC5572823
- DOI: 10.1016/j.ydbio.2016.05.038
Genetic and biochemical evidence that gastrulation defects in Pofut2 mutants result from defects in ADAMTS9 secretion
Abstract
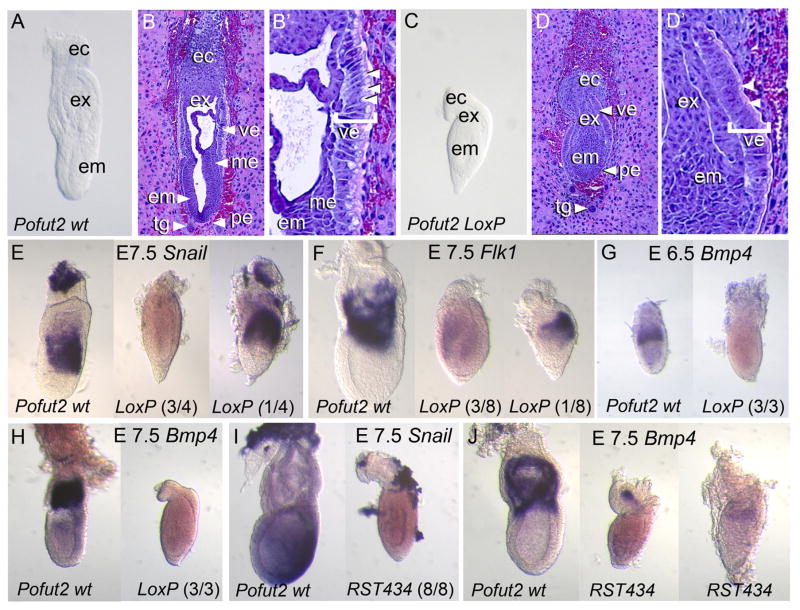
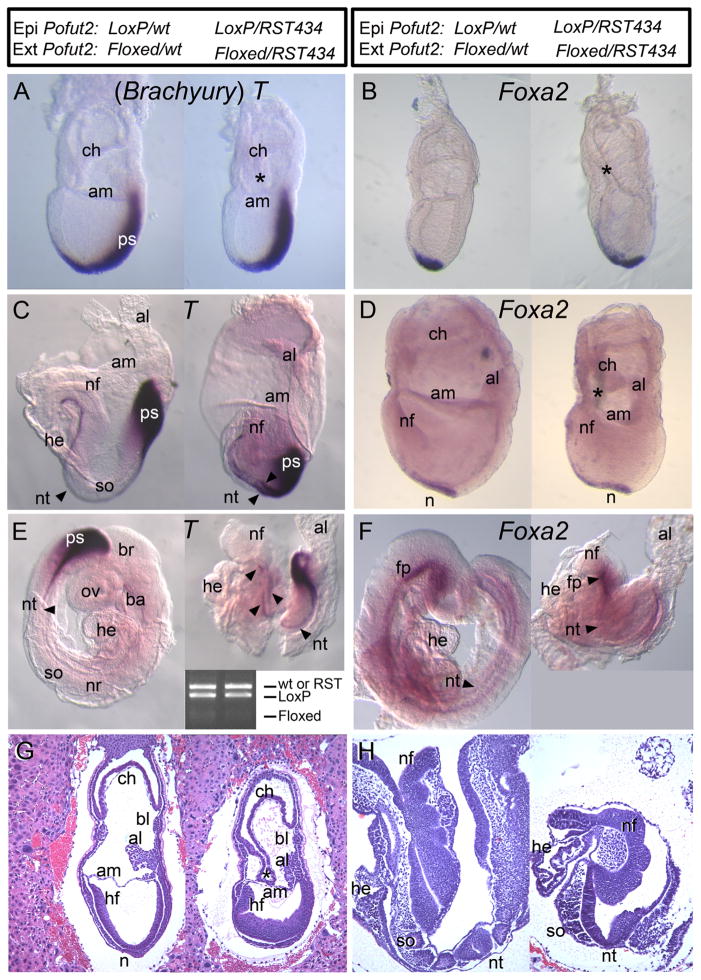
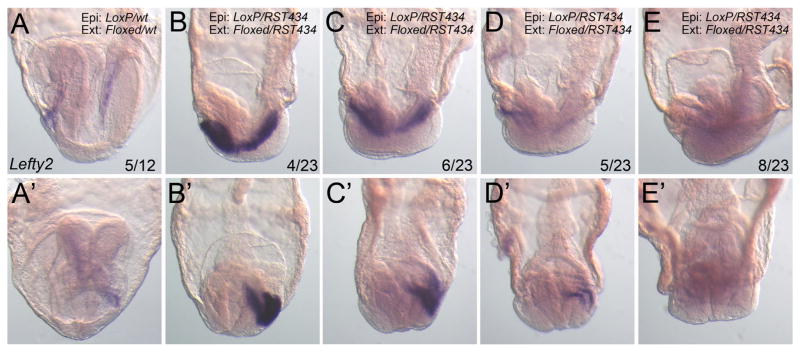
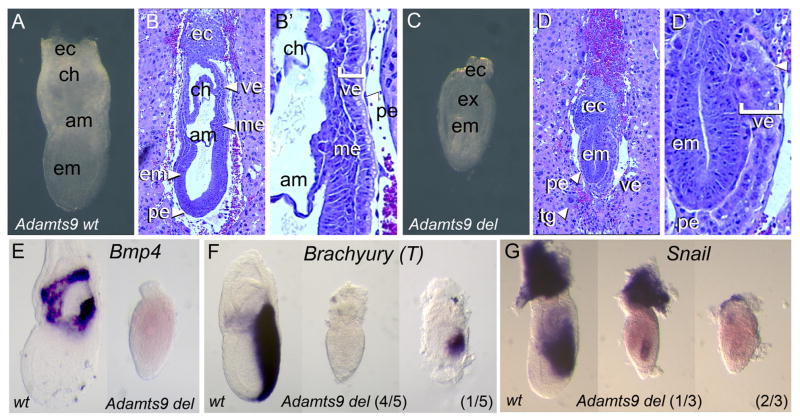
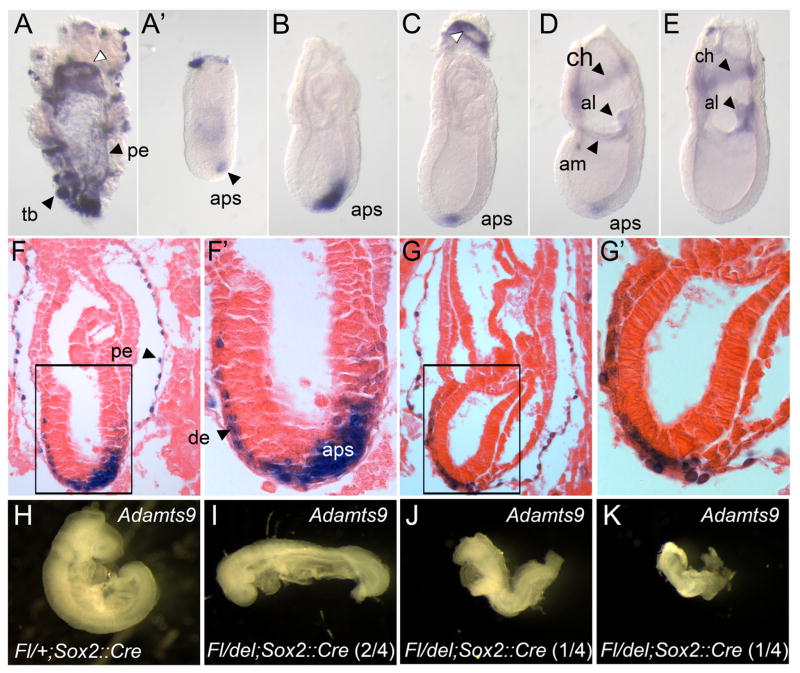
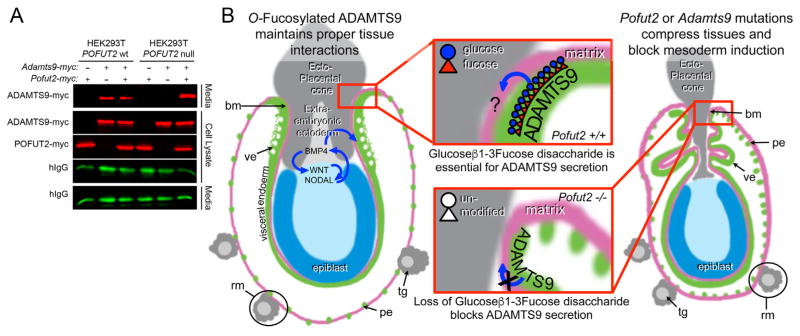
Protein O-fucosyltransferase 2 (POFUT2) adds O-linked fucose to Thrombospondin Type 1 Repeats (TSR) in 49 potential target proteins. Nearly half the POFUT2 targets belong to the A Disintegrin and Metalloprotease with ThromboSpondin type-1 motifs (ADAMTS) or ADAMTS-like family of proteins. Both the mouse Pofut2 RST434 gene trap allele and the Adamts9 knockout were reported to result in early embryonic lethality, suggesting that defects in Pofut2 mutant embryos could result from loss of O-fucosylation on ADAMTS9. To address this question, we compared the Pofut2 and Adamts9 knockout phenotypes and used Cre-mediated deletion of Pofut2 and Adamts9 to dissect the tissue-specific role of O-fucosylated ADAMTS9 during gastrulation. Disruption of Pofut2 using the knockout (LoxP) or gene trap (RST434) allele, as well as deletion of Adamts9, resulted in disorganized epithelia (epiblast, extraembryonic ectoderm, and visceral endoderm) and blocked mesoderm formation during gastrulation. The similarity between Pofut2 and Adamts9 mutants suggested that disruption of ADAMTS9 function could be responsible for the gastrulation defects observed in Pofut2 mutants. Consistent with this prediction, CRISPR/Cas9 knockout of POFUT2 in HEK293T cells blocked secretion of ADAMTS9. We determined that Adamts9 was dynamically expressed during mouse gastrulation by trophoblast giant cells, parietal endoderm, the most proximal visceral endoderm adjacent to the ectoplacental cone, extraembryonic mesoderm, and anterior primitive streak. Conditional deletion of either Pofut2 or Adamts9 in the epiblast rescues the gastrulation defects, and identified a new role for O-fucosylated ADAMTS9 during morphogenesis of the amnion and axial mesendoderm. Combined, these results suggested that loss of ADAMTS9 function in the extra embryonic tissue is responsible for gastrulation defects in the Pofut2 knockout. We hypothesize that loss of ADAMTS9 function in the most proximal visceral endoderm leads to slippage of the visceral endoderm and altered characteristics of the extraembryonic ectoderm. Consequently, loss of input from the extraembryonic ectoderm and/or compression of the epiblast by Reichert's membrane blocks gastrulation. In the future, the Pofut2 and Adamts9 knockouts will be valuable tools for understanding how local changes in the properties of the extracellular matrix influence the organization of tissues during mammalian development.
Keywords: Adamts9; Gastrulation; O-Fucosylation; Pofut2; Thrombospondin type I repeats.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Adams JC, Tucker RP. The thrombospondin type 1 repeat (TSR) superfamily: diverse proteins with related roles in neuronal development. Dev Dyn. 2000;218:280–299. - PubMed
-
- Asch AS, Silbiger S, Heimer E, Nachman RL. Thrombospondin sequence motif (CSVTCG) is responsible for CD36 binding. Biochem Biophys Res Commun. 1992;182:1208–1217. - PubMed
-
- Basu B, Brueckner M. Cilia multifunctional organelles at the center of vertebrate left-right asymmetry. Curr Top Dev Biol. 2008;85:151–174. - PubMed
-
- Beck S, Le Good JA, Guzman M, Ben Haim N, Roy K, Beermann F, Constam DB. Extraembryonic proteases regulate Nodal signalling during gastrulation. Nat Cell Biol. 2002;4:981–985. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
