

Divergent regulation of functionally distinct γ-tubulin complexes during differentiation
- PMID: 27298324
- PMCID: PMC4915192
- DOI: 10.1083/jcb.201601099
Divergent regulation of functionally distinct γ-tubulin complexes during differentiation
Abstract
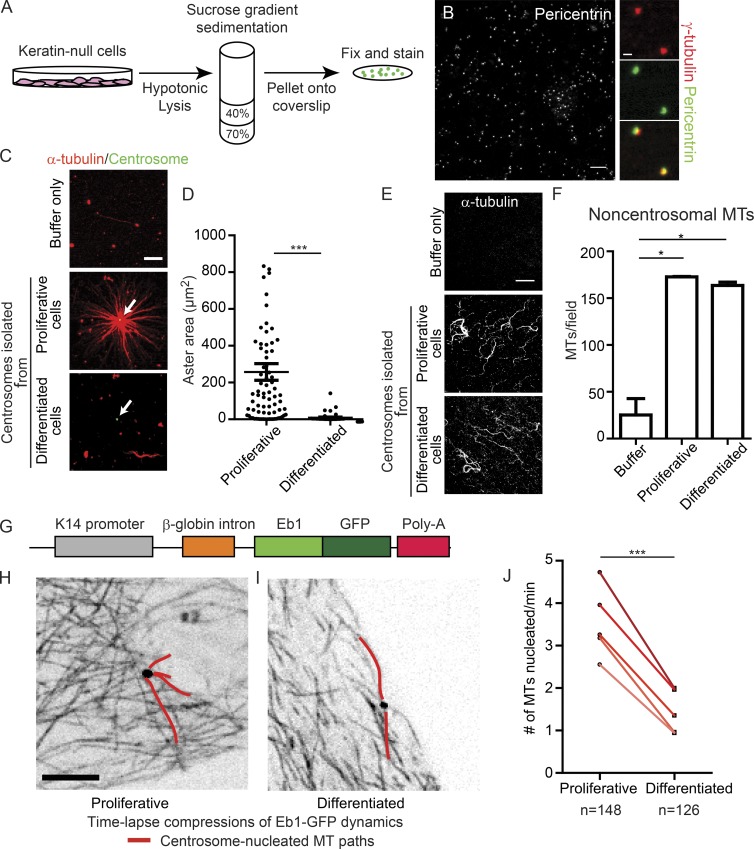
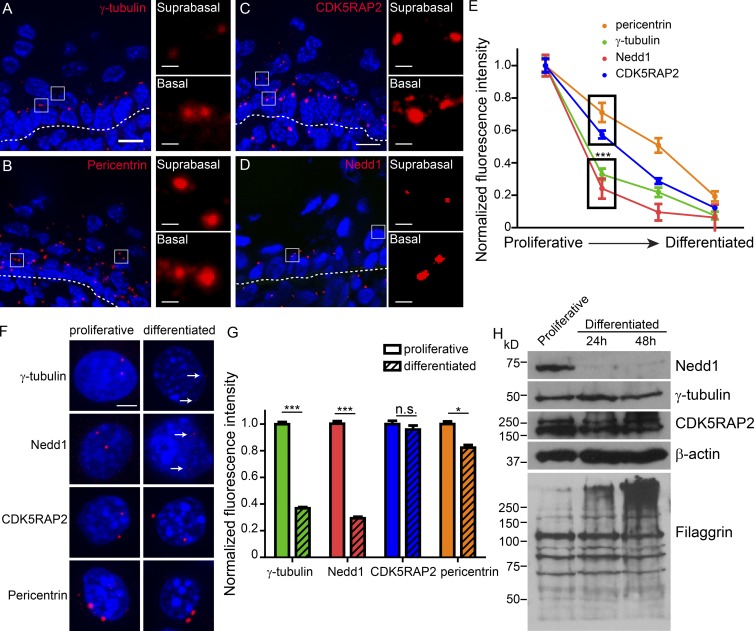
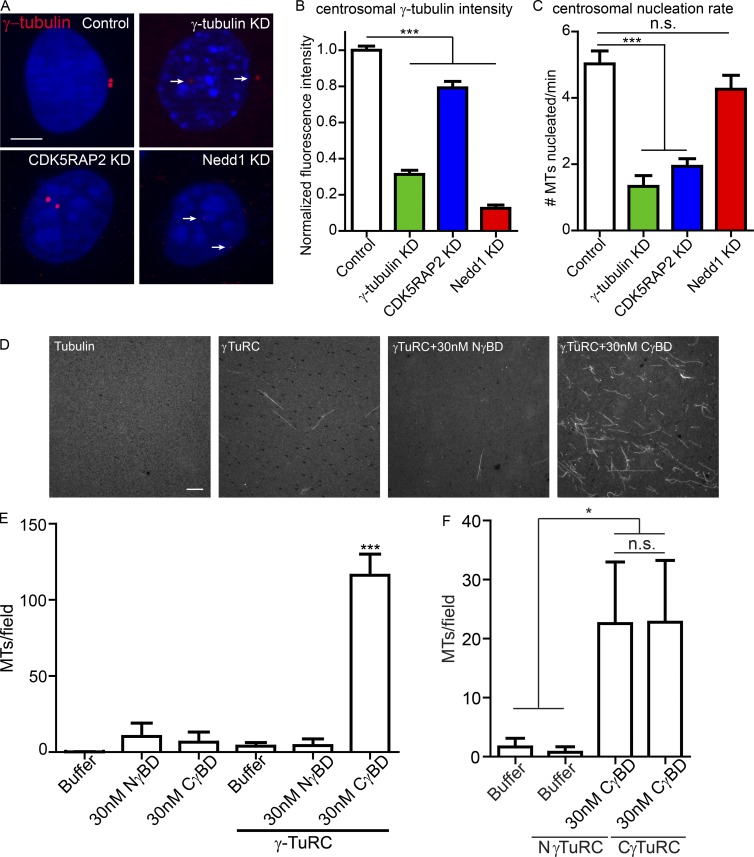
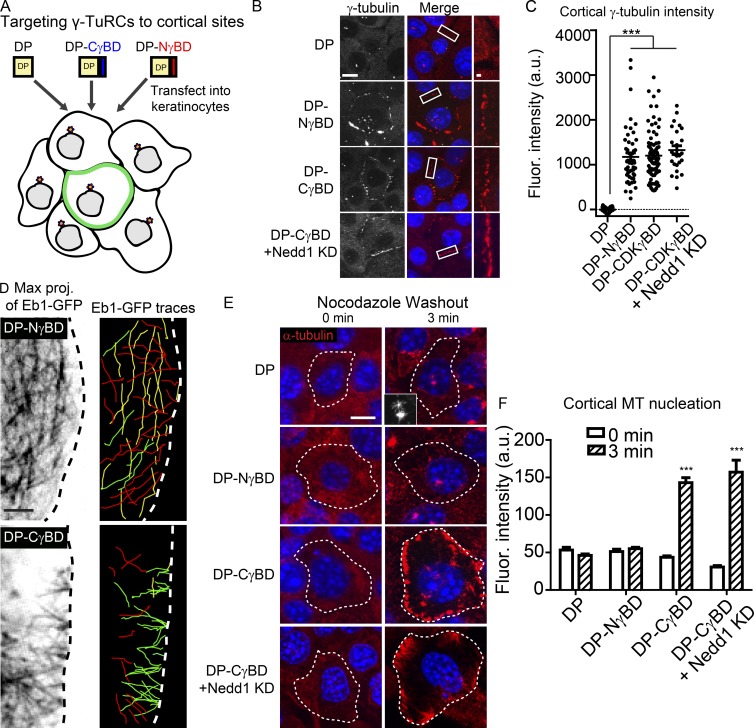
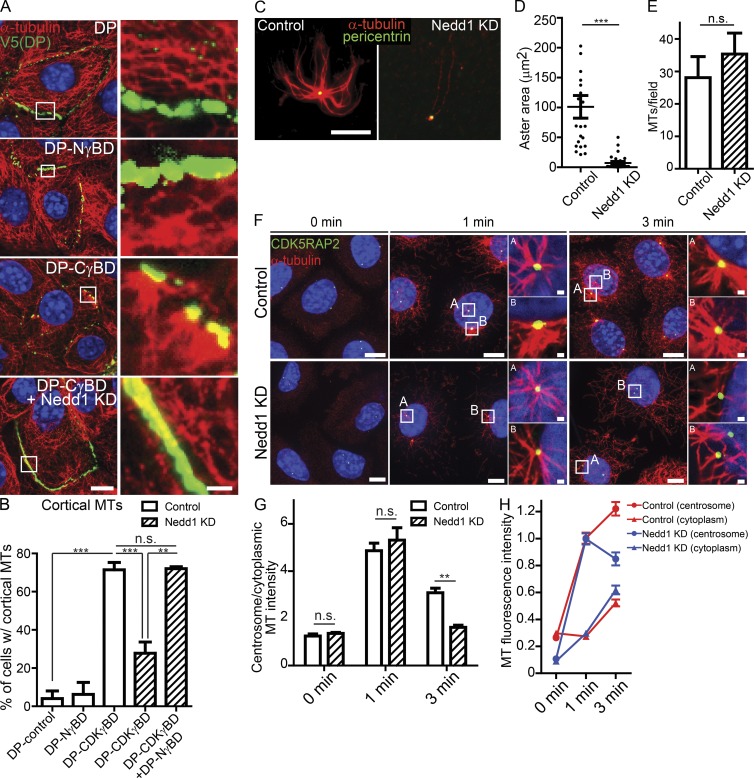
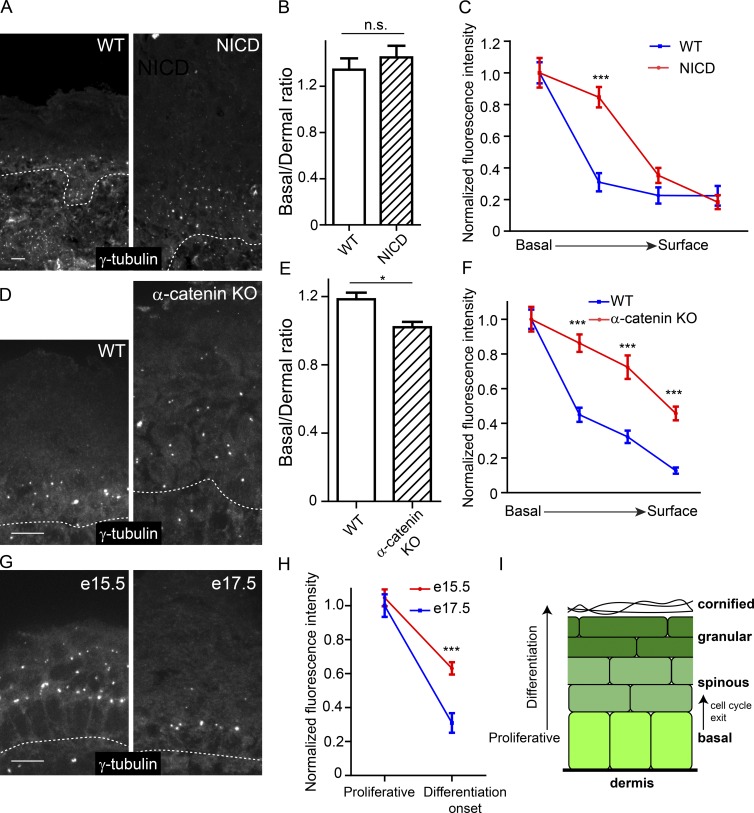
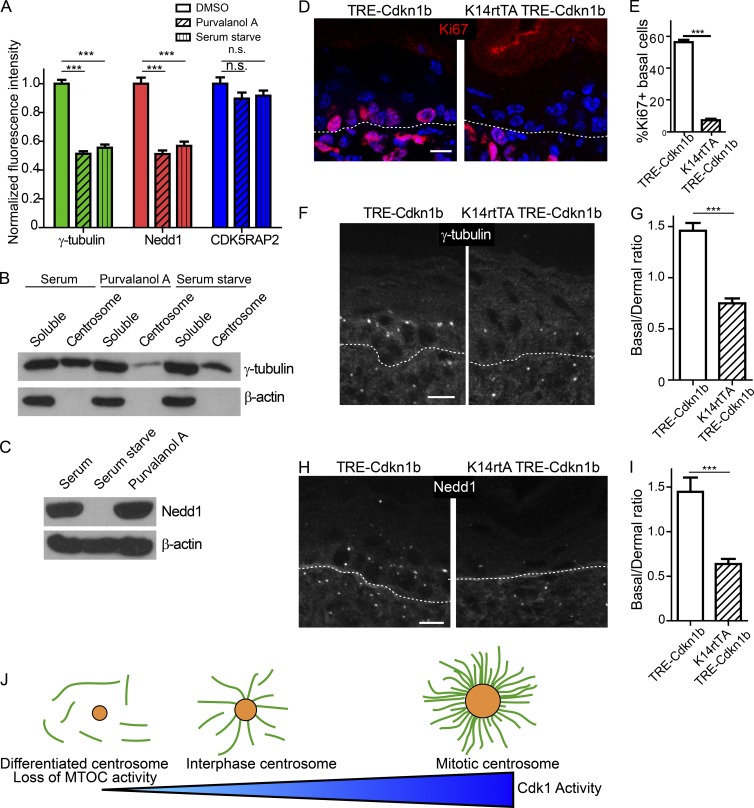
Differentiation induces the formation of noncentrosomal microtubule arrays in diverse tissues. The formation of these arrays requires loss of microtubule-organizing activity (MTOC) at the centrosome, but the mechanisms regulating this transition remain largely unexplored. Here, we use the robust loss of centrosomal MTOC activity in the epidermis to identify two pools of γ-tubulin that are biochemically and functionally distinct and differentially regulated. Nucleation-competent CDK5RAP2-γ-tubulin complexes were maintained at centrosomes upon initial epidermal differentiation. In contrast, Nedd1-γ-tubulin complexes did not promote nucleation but were required for anchoring of microtubules, a previously uncharacterized activity for this complex. Cell cycle exit specifically triggered loss of Nedd1-γ-tubulin complexes, providing a mechanistic link connecting MTOC activity and differentiation. Collectively, our studies demonstrate that distinct γ-tubulin complexes regulate different microtubule behaviors at the centrosome and show that differential regulation of these complexes drives loss of centrosomal MTOC activity.
© 2016 Muroyama et al.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
