

Mechanisms of Intestinal Barrier Dysfunction in Sepsis
- PMID: 27299587
- PMCID: PMC4910519
- DOI: 10.1097/SHK.0000000000000565
Mechanisms of Intestinal Barrier Dysfunction in Sepsis
Abstract
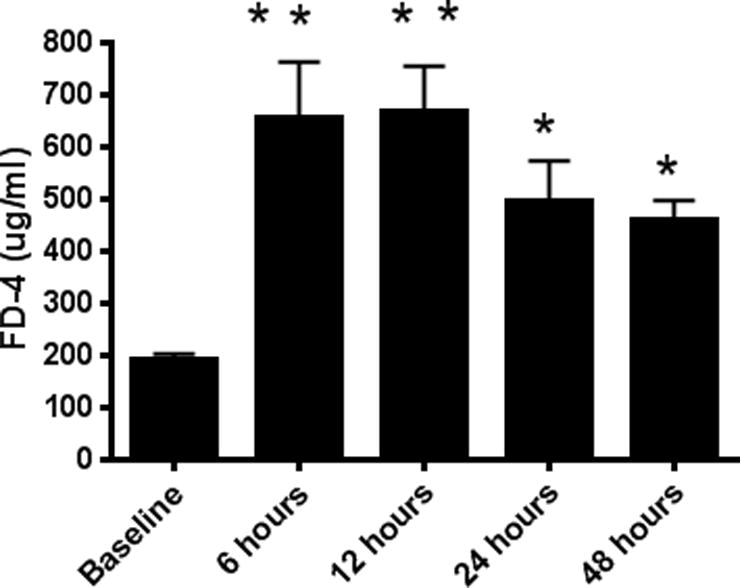
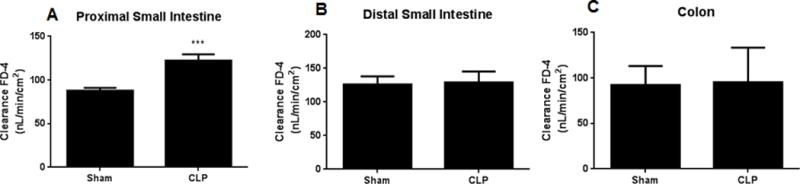
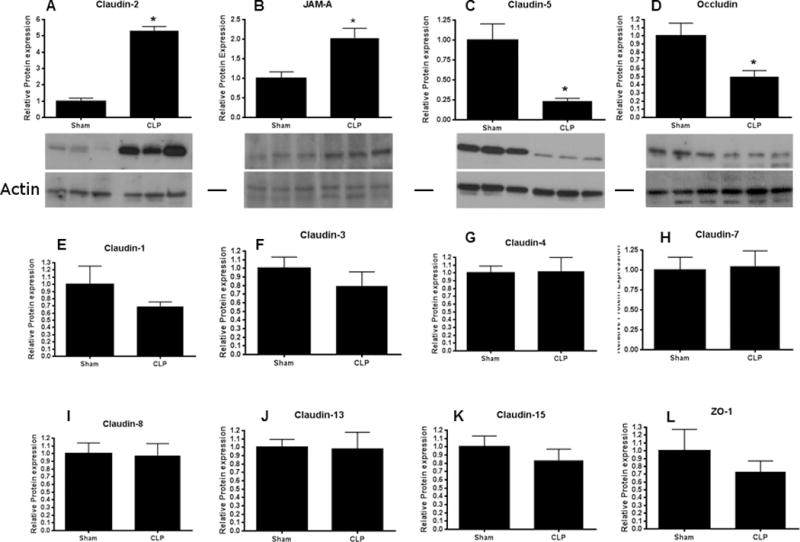
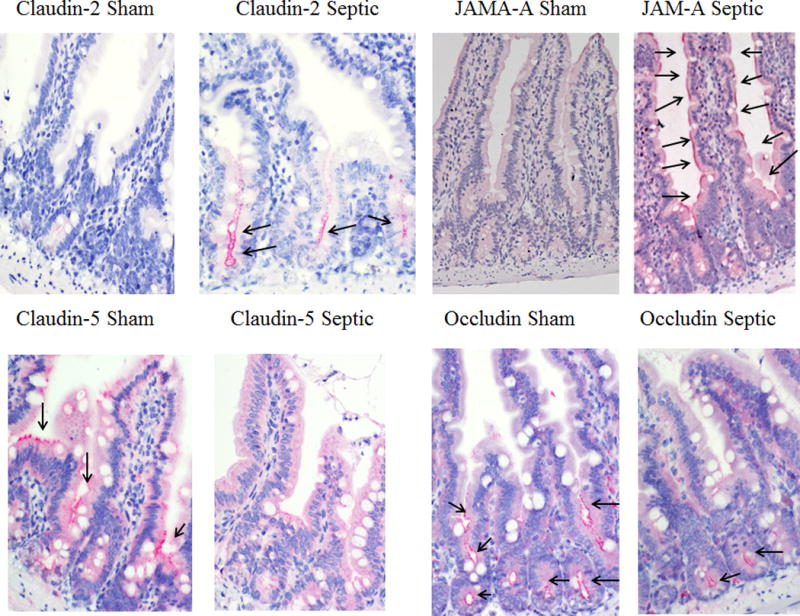
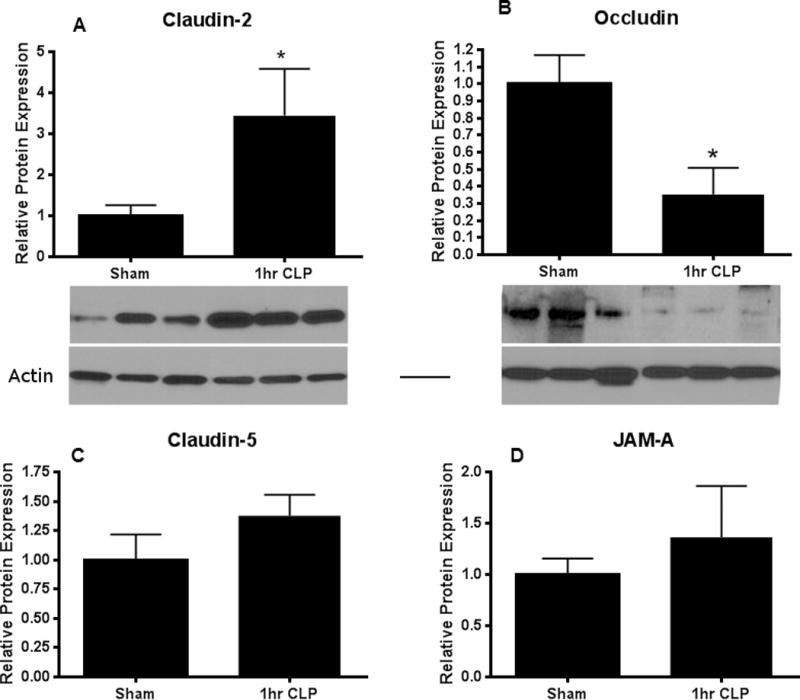
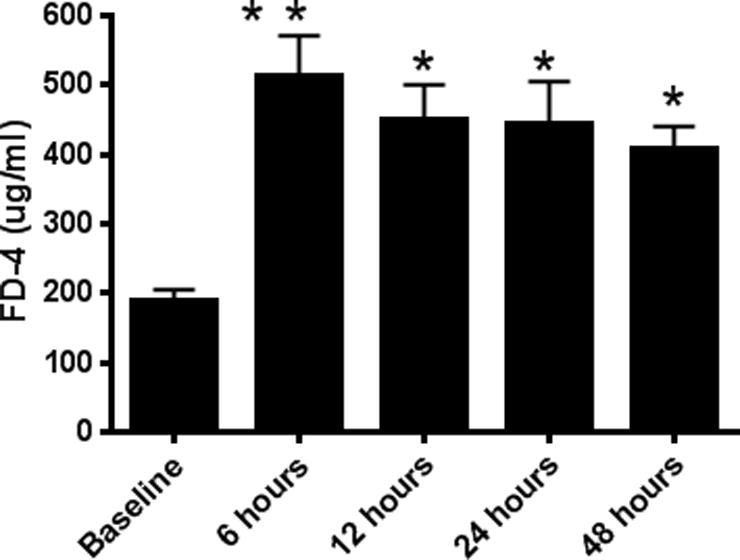
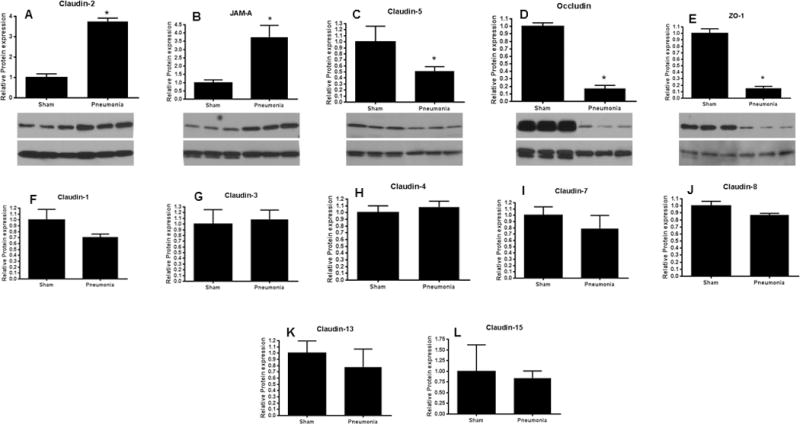
Intestinal barrier dysfunction is thought to contribute to the development of multiple organ dysfunction syndrome in sepsis. Although there are similarities in clinical course following sepsis, there are significant differences in the host response depending on the initiating organism and time course of the disease, and pathways of gut injury vary widely in different preclinical models of sepsis. The purpose of this study was to determine whether the timecourse and mechanisms of intestinal barrier dysfunction are similar in disparate mouse models of sepsis with similar mortalities. FVB/N mice were randomized to receive cecal ligation and puncture (CLP) or sham laparotomy, and permeability was measured to fluoresceinisothiocyanate conjugated-dextran (FD-4) six to 48 h later. Intestinal permeability was elevated following CLP at all timepoints measured, peaking at 6 to 12 h. Tight junction proteins claudin 1, 2, 3, 4, 5, 7, 8, 13, and 15, Junctional Adhesion Molecule-A (JAM-A), occludin, and ZO-1 were than assayed by Western blot, real-time polymerase chain reaction, and immunohistochemistry 12 h after CLP to determine potential mechanisms underlying increases in intestinal permeability. Claudin 2 and JAM-A were increased by sepsis, whereas claudin-5 and occludin were decreased by sepsis. All other tight junction proteins were unchanged. A further timecourse experiment demonstrated that alterations in claudin-2 and occludin were detectable as early as 1 h after the onset of sepsis. Similar experiments were then performed in a different group of mice subjected to Pseudomonas aeruginosa pneumonia. Mice with pneumonia had an increase in intestinal permeability similar in timecourse and magnitude to that seen in CLP. Similar changes in tight junction proteins were seen in both models of sepsis although mice subjected to pneumonia also had a marked decrease in ZO-1 not seen in CLP. These results indicate that two disparate, clinically relevant models of sepsis induce a significant increase in intestinal permeability mediated through a common pathway involving alterations in claudin 2, claudin 5, JAM-A, and occludin although model-specific differences in ZO-1 were also identified.
Conflict of interest statement
Conflicts of Interest: Craig Coopersmith was the President of the Society of Critical Care Medicine when this work was submitted.
Figures
References
-
- Gaieski DF, Edwards JM, Kallan MJ, Carr BG. Benchmarking the incidence and mortality of severe sepsis in the United States. Crit Care Med. 2013;41:1167–1174. - PubMed
-
- Levy MM, Rhodes A, Phillips GS, Townsend SR, Schorr CA, Beale R, Osborn T, Lemeshow S, Chiche JD, Artigas A, Dellinger RP. Surviving sepsis campaign: association between performance metrics and outcomes in a 7.5-year study. Crit Care Med. 2015;43:3–12. - PubMed
-
- McConnell KW, McDunn JE, Clark AT, Dunne WM, Dixon DJ, Turnbull IR, Dipasco PJ, Osberghaus WF, Sherman B, Martin JR, Walter MJ, Cobb JP, Buchman TG, Hotchkiss RS, Coopersmith CM. Streptococcus pneumoniae and Pseudomonas aeruginosa pneumonia induce distinct host responses. Crit Care Med. 2010;38:223–241. - PMC - PubMed
-
- Fry DE. The generic septic response. Crit Care Med. 2008;36:1369–1370. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
