

Arabidopsis late blight: infection of a nonhost plant by Albugo laibachii enables full colonization by Phytophthora infestans
- PMID: 27302335
- PMCID: PMC5215655
- DOI: 10.1111/cmi.12628
Arabidopsis late blight: infection of a nonhost plant by Albugo laibachii enables full colonization by Phytophthora infestans
Abstract
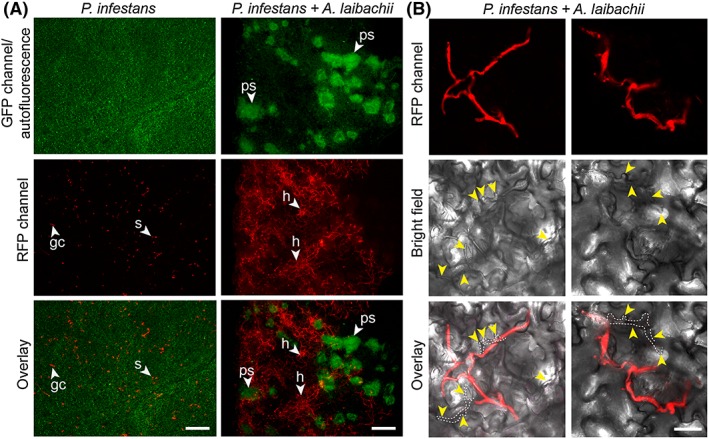
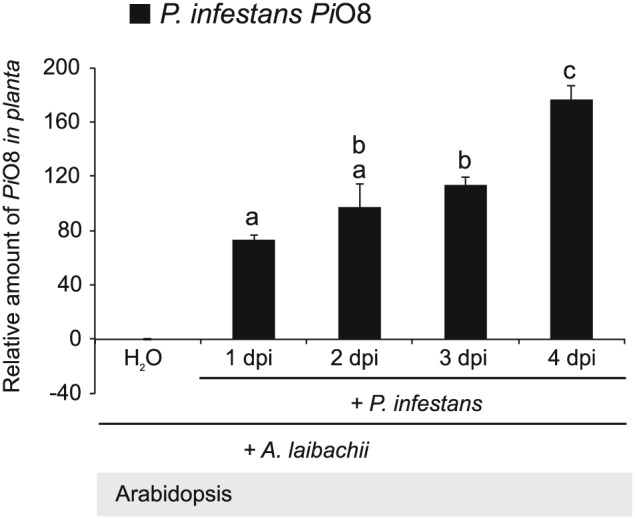
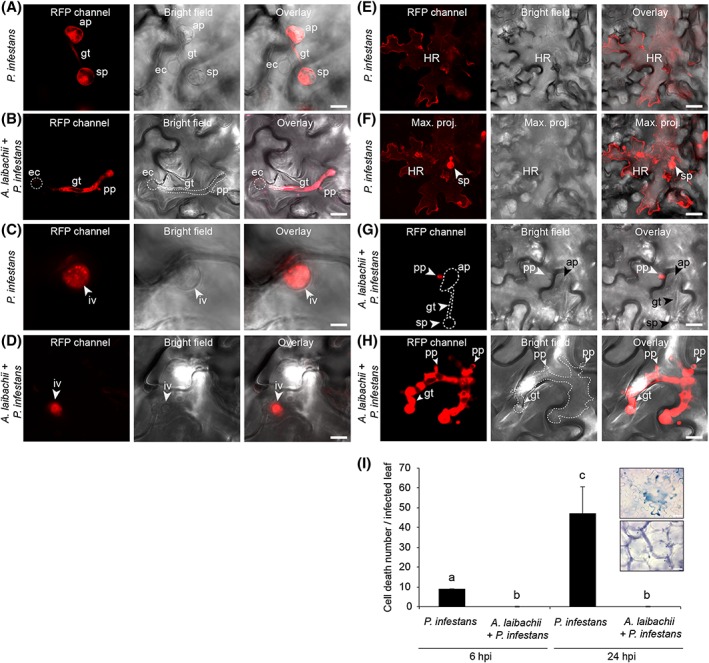
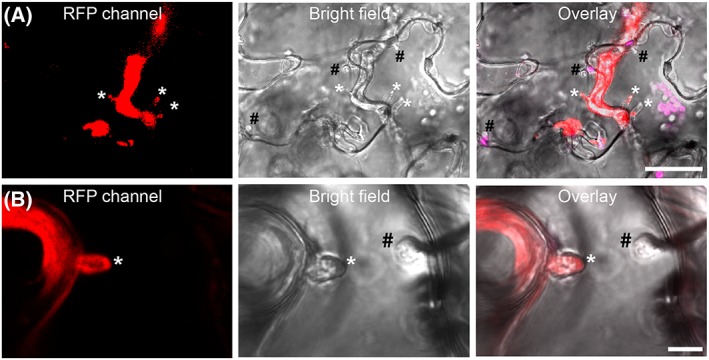
The oomycete pathogen Phytophthora infestans causes potato late blight, and as a potato and tomato specialist pathogen, is seemingly poorly adapted to infect plants outside the Solanaceae. Here, we report the unexpected finding that P. infestans can infect Arabidopsis thaliana when another oomycete pathogen, Albugo laibachii, has colonized the host plant. The behaviour and speed of P. infestans infection in Arabidopsis pre-infected with A. laibachii resemble P. infestans infection of susceptible potato plants. Transcriptional profiling of P. infestans genes during infection revealed a significant overlap in the sets of secreted-protein genes that are induced in P. infestans upon colonization of potato and susceptible Arabidopsis, suggesting major similarities in P. infestans gene expression dynamics on the two plant species. Furthermore, we found haustoria of A. laibachii and P. infestans within the same Arabidopsis cells. This Arabidopsis-A. laibachii-P. infestans tripartite interaction opens up various possibilities to dissect the molecular mechanisms of P. infestans infection and the processes occurring in co-infected Arabidopsis cells.
© 2016 The Authors Cellular Microbiology Published by John Wiley & Sons Ltd.
Figures


References
-
- Agrios, G.N. (2005) Plant Pathology. San Diego, CA: Elsevier.
-
- Akrofi, A.Y. , Amoako‐Atta, I. , Assuah, M. , and Asare, E.K. (2015) Black pod disease on cacao (Theobroma cacao, L) in Ghana: spread of Phytophthora megakarya and role of economic plants in the disease epidemiology. Crop Prot 72: 66–75.
-
- Ayliffe, M. , Jin, Y. , Kang, Z. , Persson, M. , Steffenson, B. , Wang, S. , and Leung, H. (2011) Determining the basis of nonhost resistance in rice to cereal rusts. Euphytica 179: 33–40.
-
- Bains, S. , and Jhooty, J. (1985) Association of Peronospora parasitica with Albugo candida on Brassica juncea leaves. J Phytopathol 112: 28–31.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
