

Extracellular matrix motion and early morphogenesis
- PMID: 27302396
- PMCID: PMC4920166
- DOI: 10.1242/dev.127886
Extracellular matrix motion and early morphogenesis
Abstract
For over a century, embryologists who studied cellular motion in early amniotes generally assumed that morphogenetic movement reflected migration relative to a static extracellular matrix (ECM) scaffold. However, as we discuss in this Review, recent investigations reveal that the ECM is also moving during morphogenesis. Time-lapse studies show how convective tissue displacement patterns, as visualized by ECM markers, contribute to morphogenesis and organogenesis. Computational image analysis distinguishes between cell-autonomous (active) displacements and convection caused by large-scale (composite) tissue movements. Modern quantification of large-scale 'total' cellular motion and the accompanying ECM motion in the embryo demonstrates that a dynamic ECM is required for generation of the emergent motion patterns that drive amniote morphogenesis.
Keywords: Amniote morphogenesis; Emergent patterns; Extracellular matrix dynamics; Tissue-scale motion.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
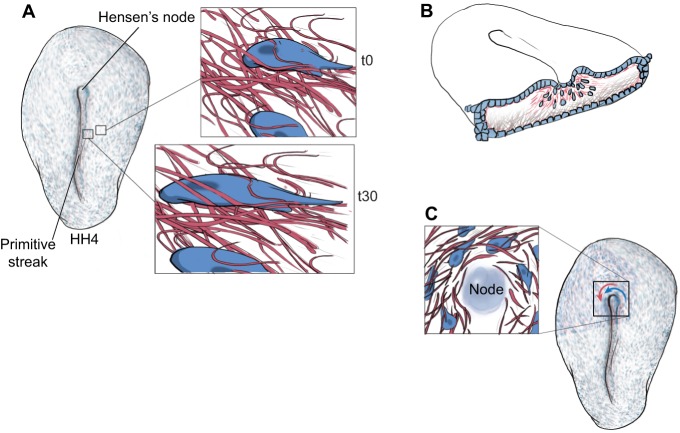
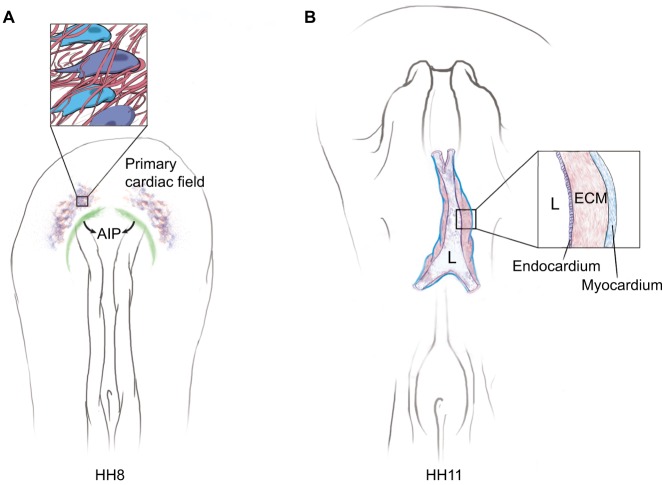
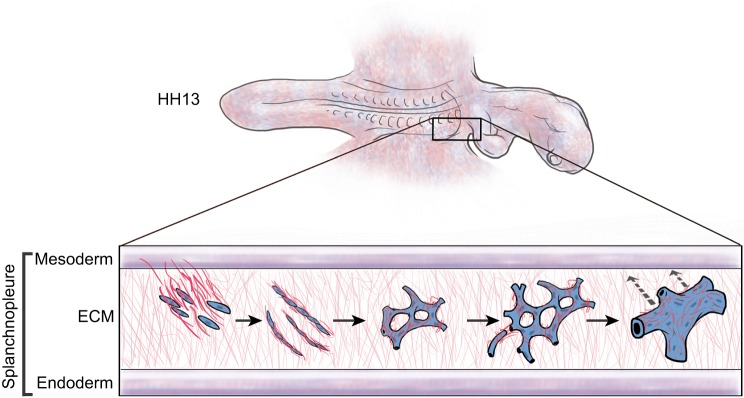
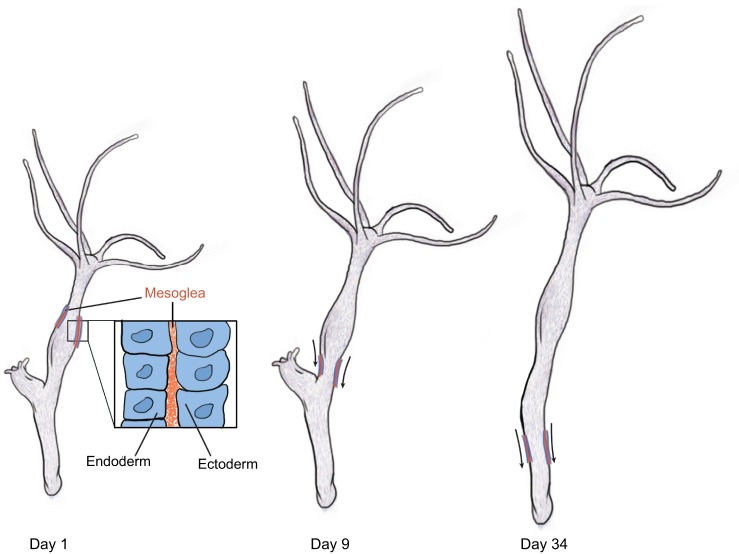
References
-
- Adams J. C. (2013). Extracellular matrix evolution: an overview. In Evolution of Extracellular Matrix (ed. Keeley F. W. and Mecham R. P.), pp. 1-25. Berlin: Springer.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
