

TgATAT-Mediated α-Tubulin Acetylation Is Required for Division of the Protozoan Parasite Toxoplasma gondii
- PMID: 27303695
- PMCID: PMC4863603
- DOI: 10.1128/mSphere.00088-15
TgATAT-Mediated α-Tubulin Acetylation Is Required for Division of the Protozoan Parasite Toxoplasma gondii
Abstract
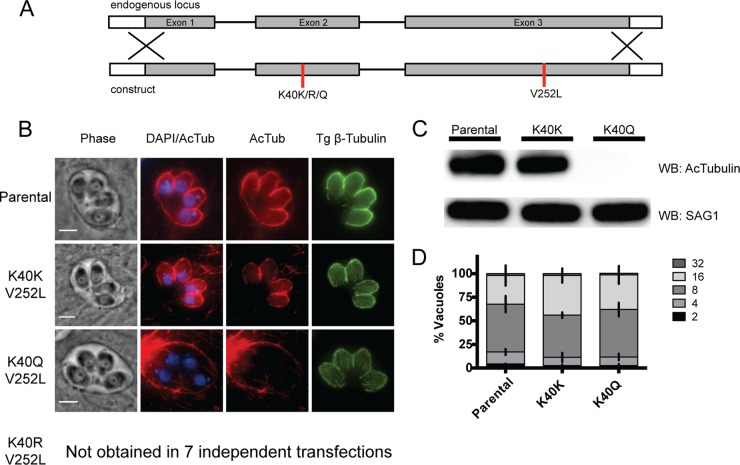
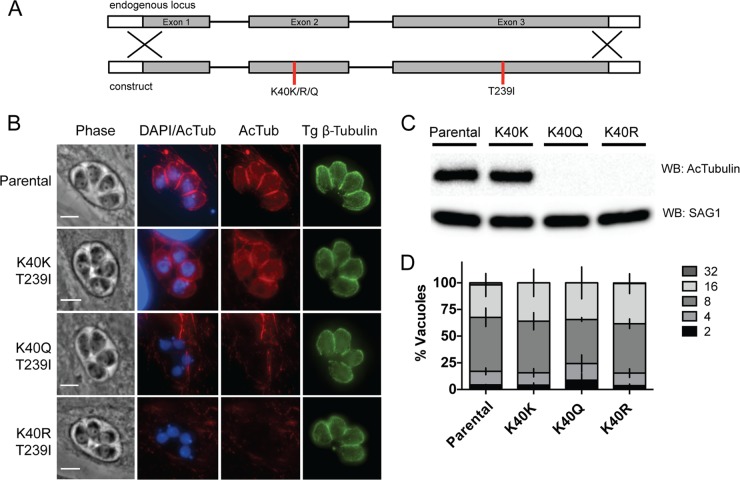
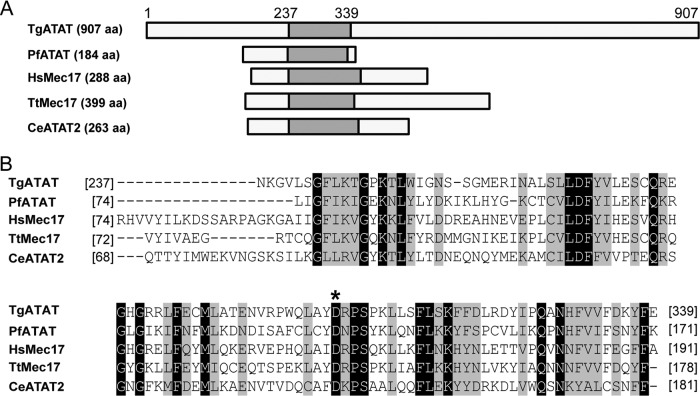
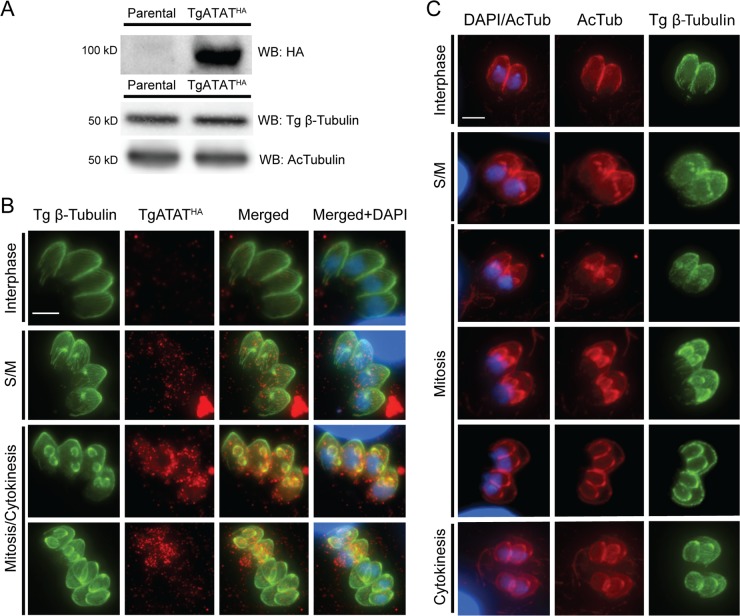
Toxoplasma gondii is a widespread protozoan parasite that causes potentially life-threatening opportunistic disease. New inhibitors of parasite replication are urgently needed, as the current antifolate treatment is also toxic to patients. Microtubules are essential cytoskeletal components that have been selectively targeted in microbial pathogens; further study of tubulin in Toxoplasma may reveal novel therapeutic opportunities. It has been noted that α-tubulin acetylation at lysine 40 (K40) is enriched during daughter parasite formation, but the impact of this modification on Toxoplasma division and the enzyme mediating its delivery have not been identified. We performed mutational analyses to provide evidence that K40 acetylation stabilizes Toxoplasma microtubules and is required for parasite replication. We also show that an unusual Toxoplasma homologue of α-tubulin acetyltransferase (TgATAT) is expressed in a cell cycle-regulated manner and that its expression peaks during division. Disruption of TgATAT with CRISPR/Cas9 ablates K40 acetylation and induces replication defects; parasites appear to initiate mitosis yet exhibit incomplete or improper nuclear division. Together, these findings establish the importance of tubulin acetylation, exposing a new vulnerability in Toxoplasma that could be pharmacologically targeted. IMPORTANCE Toxoplasma gondii is an opportunistic parasite that infects at least one-third of the world population. New treatments for the disease (toxoplasmosis) are needed since current drugs are toxic to patients. Microtubules are essential cellular structures built from tubulin that show promise as antimicrobial drug targets. Microtubules can be regulated by chemical modification, such as acetylation on lysine 40 (K40). To determine the role of K40 acetylation in Toxoplasma and whether it is a liability to the parasite, we performed mutational analyses of the α-tubulin gene. Our results indicate that parasites cannot survive without K40 acetylation unless microtubules are stabilized with a secondary mutation. Additionally, we identified the parasite enzyme that acetylates α-tubulin (TgATAT). Genetic disruption of TgATAT caused severe defects in parasite replication, further highlighting the importance of α-tubulin K40 acetylation in Toxoplasma and its promise as a potential new drug target.
Keywords: Mec-17; acetyltransferase; cytoskeleton; endodyogeny; lysine acetylation; microtubules.
Figures
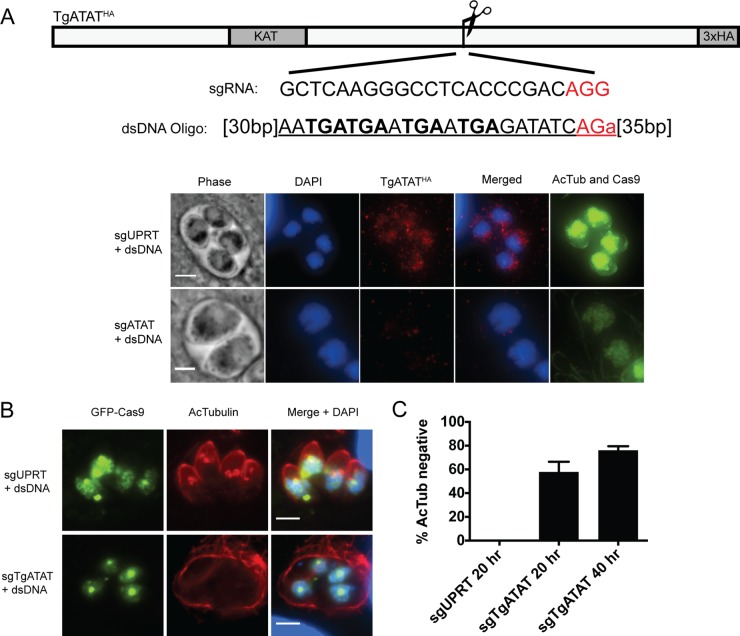
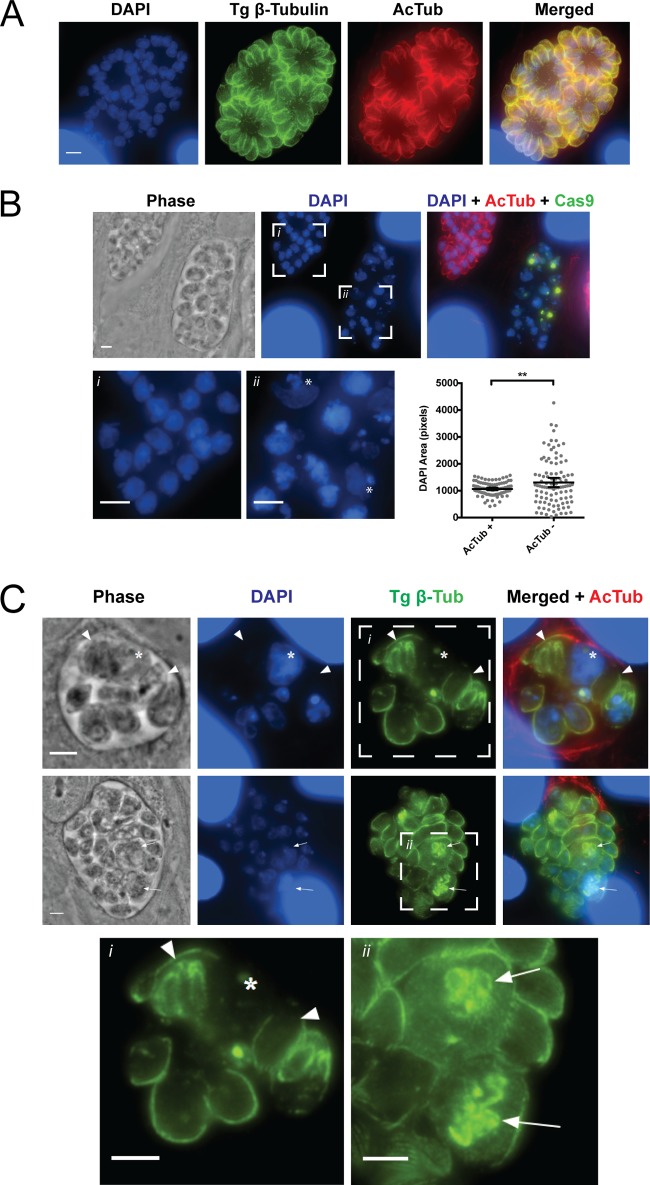
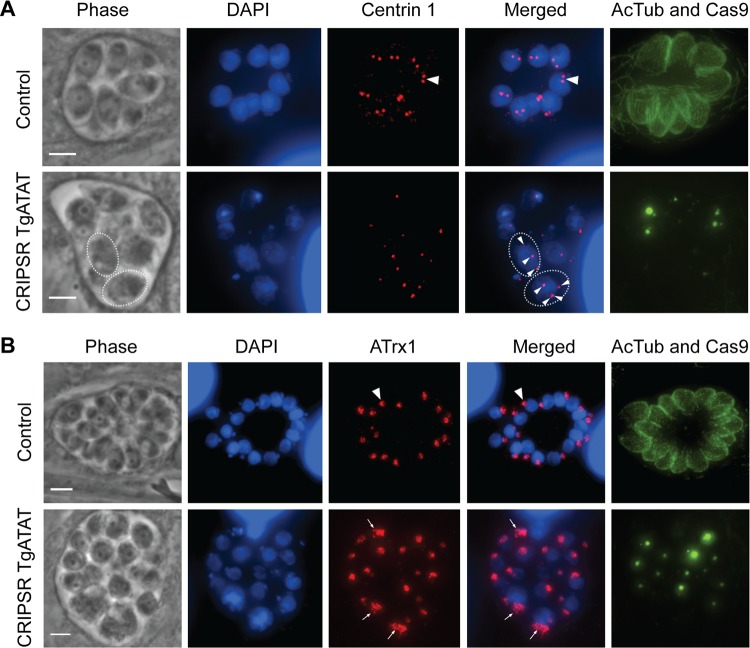
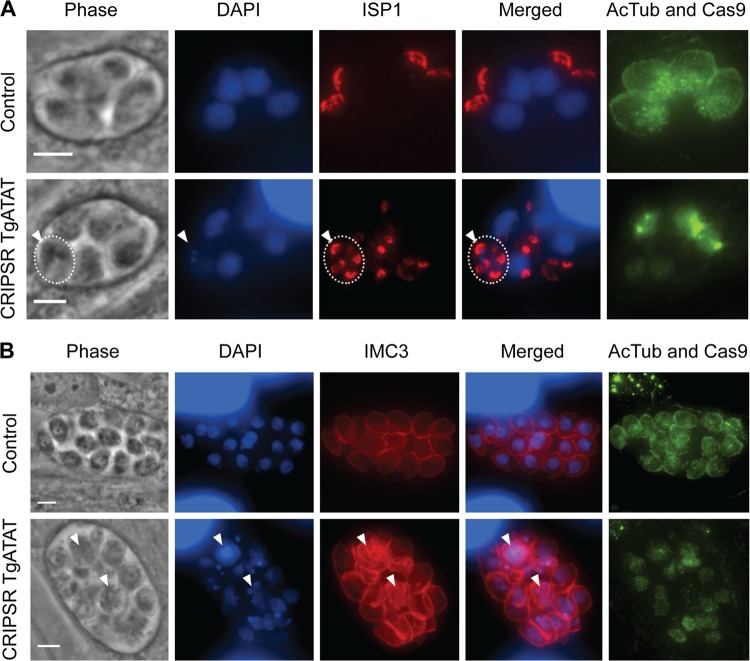
Similar articles
-
Alpha-Tubulin Acetylation in Trypanosoma cruzi: A Dynamic Instability of Microtubules Is Required for Replication and Cell Cycle Progression.Front Cell Infect Microbiol. 2021 Mar 11;11:642271. doi: 10.3389/fcimb.2021.642271. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 33777851 Free PMC article.
-
Lysine acetylation is widespread on proteins of diverse function and localization in the protozoan parasite Toxoplasma gondii.Eukaryot Cell. 2012 Jun;11(6):735-42. doi: 10.1128/EC.00088-12. Epub 2012 Apr 27. Eukaryot Cell. 2012. PMID: 22544907 Free PMC article.
-
MEC-17 is an alpha-tubulin acetyltransferase.Nature. 2010 Sep 9;467(7312):218-22. doi: 10.1038/nature09324. Nature. 2010. PMID: 20829795 Free PMC article.
-
Targeting Toxoplasma tubules: tubulin, microtubules, and associated proteins in a human pathogen.Eukaryot Cell. 2015 Jan;14(1):2-12. doi: 10.1128/EC.00225-14. Epub 2014 Nov 7. Eukaryot Cell. 2015. PMID: 25380753 Free PMC article. Review.
-
The growing landscape of tubulin acetylation: lysine 40 and many more.Biochem J. 2016 Jul 1;473(13):1859-68. doi: 10.1042/BCJ20160172. Biochem J. 2016. PMID: 27354562 Review.
Cited by
-
A novel dense granule protein, GRA41, regulates timing of egress and calcium sensitivity in Toxoplasma gondii.Cell Microbiol. 2017 Sep;19(9):10.1111/cmi.12749. doi: 10.1111/cmi.12749. Epub 2017 May 17. Cell Microbiol. 2017. PMID: 28436089 Free PMC article.
-
Characterization of Plasmodium Atg3-Atg8 Interaction Inhibitors Identifies Novel Alternative Mechanisms of Action in Toxoplasma gondii.Antimicrob Agents Chemother. 2018 Jan 25;62(2):e01489-17. doi: 10.1128/AAC.01489-17. Print 2018 Feb. Antimicrob Agents Chemother. 2018. PMID: 29158278 Free PMC article.
-
Alpha-Tubulin Acetylation in Trypanosoma cruzi: A Dynamic Instability of Microtubules Is Required for Replication and Cell Cycle Progression.Front Cell Infect Microbiol. 2021 Mar 11;11:642271. doi: 10.3389/fcimb.2021.642271. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 33777851 Free PMC article.
-
Tubulin Post-Translational Modifications: The Elusive Roles of Acetylation.Biology (Basel). 2023 Apr 6;12(4):561. doi: 10.3390/biology12040561. Biology (Basel). 2023. PMID: 37106761 Free PMC article. Review.
-
QTL Mapping and CRISPR/Cas9 Editing to Identify a Drug Resistance Gene in Toxoplasma gondii.J Vis Exp. 2017 Jun 22;(124):55185. doi: 10.3791/55185. J Vis Exp. 2017. PMID: 28671645 Free PMC article.
References
-
- Luft BJ, Brooks RG, Conley FK, McCabe RE, Remington JS. 1984. Toxoplasmic encephalitis in patients with acquired immune deficiency syndrome. JAMA 252:913–917. - PubMed
-
- Nissapatorn V, Lee C, Quek KF, Leong CL, Mahmud R, Abdullah KA. 2004. Toxoplasmosis in HIV/AIDS patients: a current situation. Jpn J Infect Dis 57:160–165. - PubMed
-
- Robert-Gangneux F, Sterkers Y, Yera H, Accoceberry I, Menotti J, Cassaing S, Brenier-Pinchart MP, Hennequin C, Delhaes L, Bonhomme J, Villena I, Scherer E, Dalle F, Touafek F, Filisetti D, Varlet-Marie E, Pelloux H, Bastien P. 2015. Molecular diagnosis of toxoplasmosis in immunocompromised patients: a 3-year multicenter retrospective study. J Clin Microbiol 53:1677–1684. doi:10.1128/JCM.03282-14. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
