

Effects of Sex, Strain, and Energy Intake on Hallmarks of Aging in Mice
- PMID: 27304509
- PMCID: PMC4911707
- DOI: 10.1016/j.cmet.2016.05.027
Effects of Sex, Strain, and Energy Intake on Hallmarks of Aging in Mice
Abstract
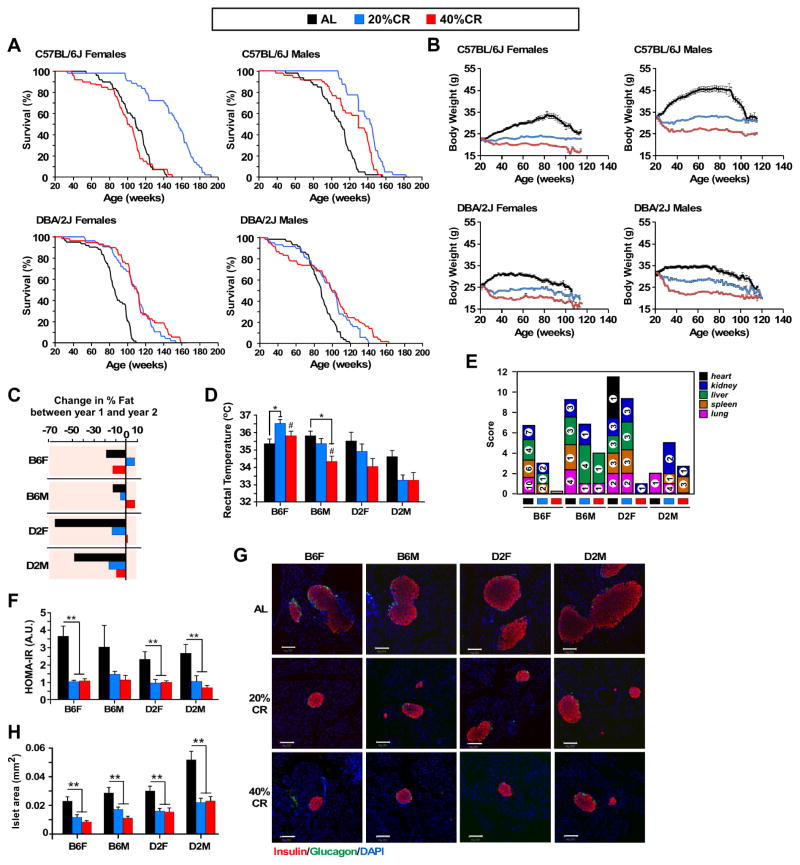
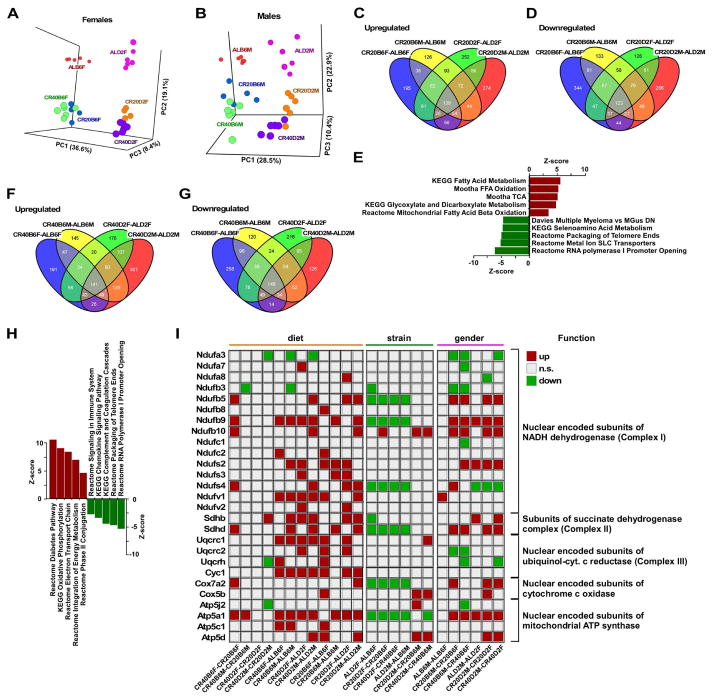
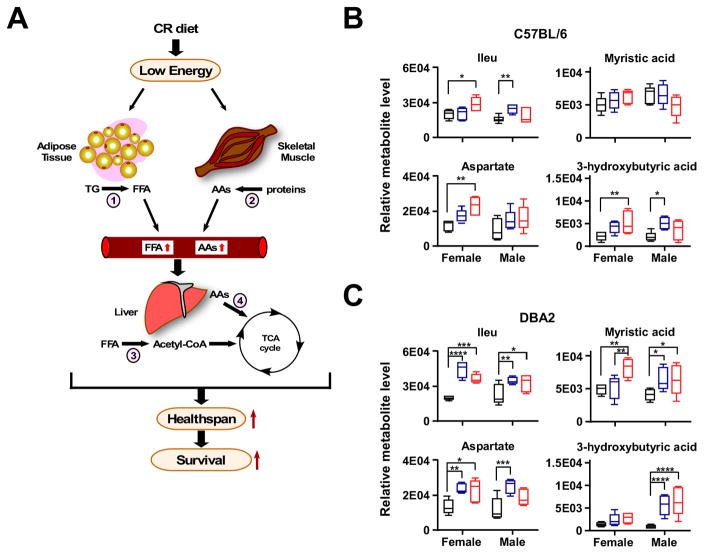
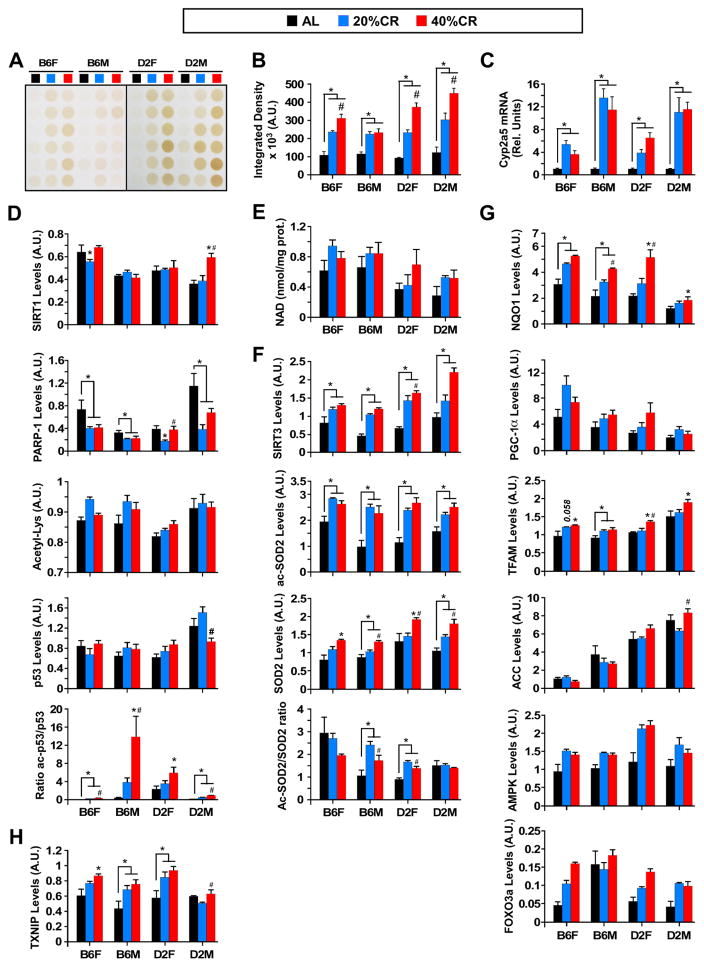
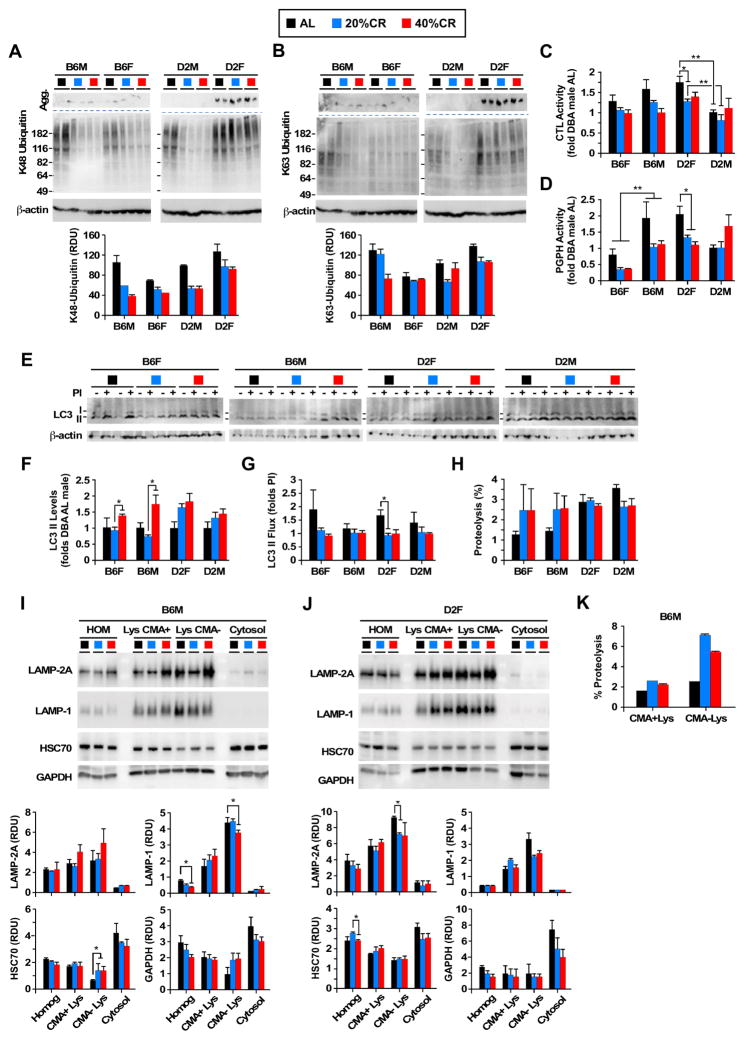
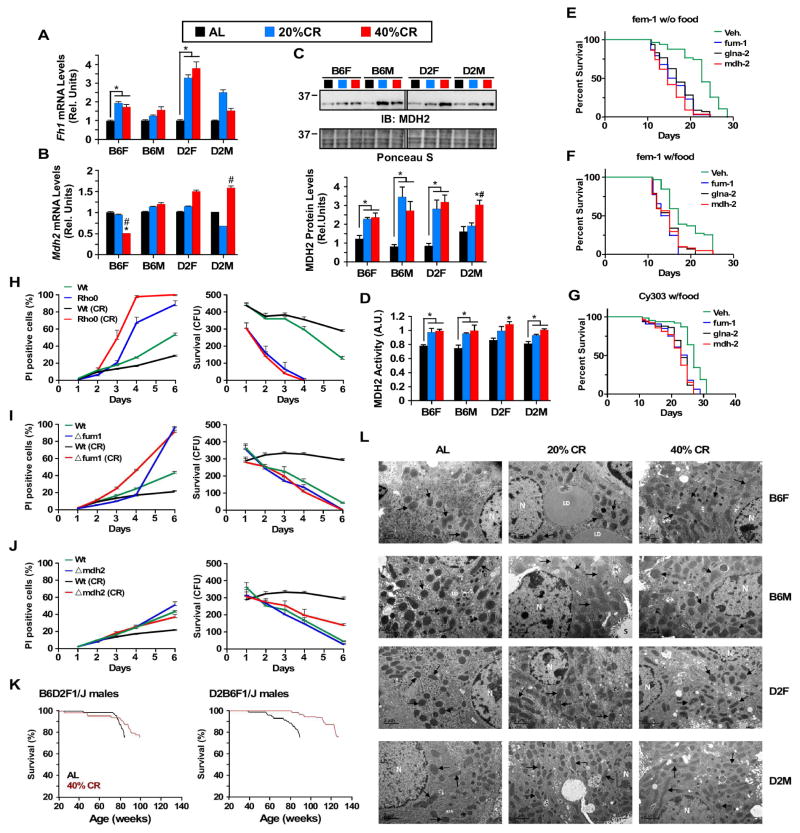
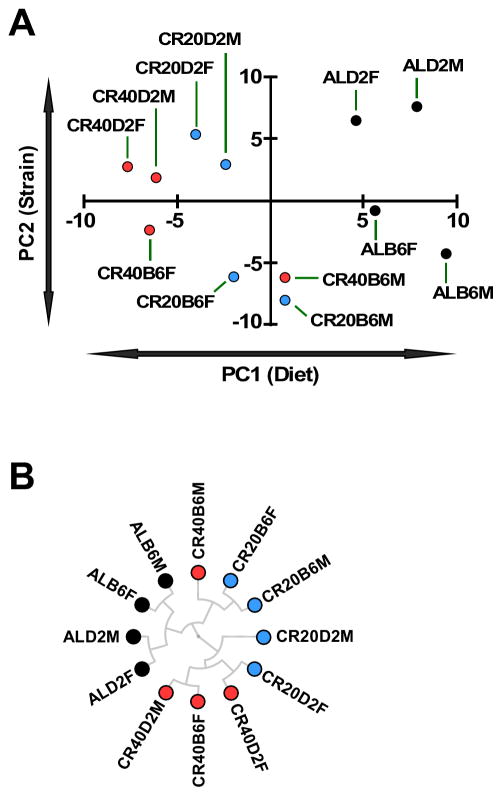
Calorie restriction (CR) is the most robust non-genetic intervention to delay aging. However, there are a number of emerging experimental variables that alter CR responses. We investigated the role of sex, strain, and level of CR on health and survival in mice. CR did not always correlate with lifespan extension, although it consistently improved health across strains and sexes. Transcriptional and metabolomics changes driven by CR in liver indicated anaplerotic filling of the Krebs cycle together with fatty acid fueling of mitochondria. CR prevented age-associated decline in the liver proteostasis network while increasing mitochondrial number, preserving mitochondrial ultrastructure and function with age. Abrogation of mitochondrial function negated life-prolonging effects of CR in yeast and worms. Our data illustrate the complexity of CR in the context of aging, with a clear separation of outcomes related to health and survival, highlighting complexities of translation of CR into human interventions.
Published by Elsevier Inc.
Figures
References
-
- Abu-Bakar A, Lamsa V, Arpiainen S, Moore MR, Lang MA, Hakkola J. Regulation of CYP2A5 gene by the transcription factor nuclear factor (erythroid-derived 2)-like 2. Drug Metab Dispos: the biological fate of chemicals. 2007;35:787–794. - PubMed
-
- Anisimov VN, Berstein LM, Egormin PA, Piskunova TS, Popovich IG, Zabezhinski MA, Tyndyk ML, Yurova MV, Kovalenko IG, Poroshina TE, et al. Metformin slows down aging and extends life span of female SHR mice. Cell Cycle. 2008;7:2769–2773. - PubMed
-
- Anson RM, Guo Z, de Cabo R, Iyun T, Rios M, Hagepanos A, Ingram DK, Lane MA, Mattson MP. Intermittent fasting dissociates beneficial effects of dietary restriction on glucose metabolism and neuronal resistance to injury from calorie intake. Proc Natl Acad Sci U S A. 2003;100:6216–6220. - PMC - PubMed
-
- Artal-Sanz M, Tavernarakis N. Prohibitin couples diapause signalling to mitochondrial metabolism during ageing in C. elegans. Nature. 2009;461:793–797. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 ES020812/ES/NIEHS NIH HHS/United States
- R01 DK098656/DK/NIDDK NIH HHS/United States
- R01 GM090311/GM/NIGMS NIH HHS/United States
- P30 DK041296/DK/NIDDK NIH HHS/United States
- ZIA AG000368/ImNIH/Intramural NIH HHS/United States
- P01 AG034906/AG/NIA NIH HHS/United States
- P30 ES013508/ES/NIEHS NIH HHS/United States
- P 29203/FWF_/Austrian Science Fund FWF/Austria
- P30 DK020541/DK/NIDDK NIH HHS/United States
- P30 AG038072/AG/NIA NIH HHS/United States
- P 27893/FWF_/Austrian Science Fund FWF/Austria
- P01 AG031782/AG/NIA NIH HHS/United States
- R01 AG043483/AG/NIA NIH HHS/United States
- P30 DK079626/DK/NIDDK NIH HHS/United States
- P30 AG013319/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
