

Reduced thermal sensitivity and increased opioidergic tone in the TASTPM mouse model of Alzheimer's disease
- PMID: 27306045
- PMCID: PMC5028157
- DOI: 10.1097/j.pain.0000000000000644
Reduced thermal sensitivity and increased opioidergic tone in the TASTPM mouse model of Alzheimer's disease
Abstract
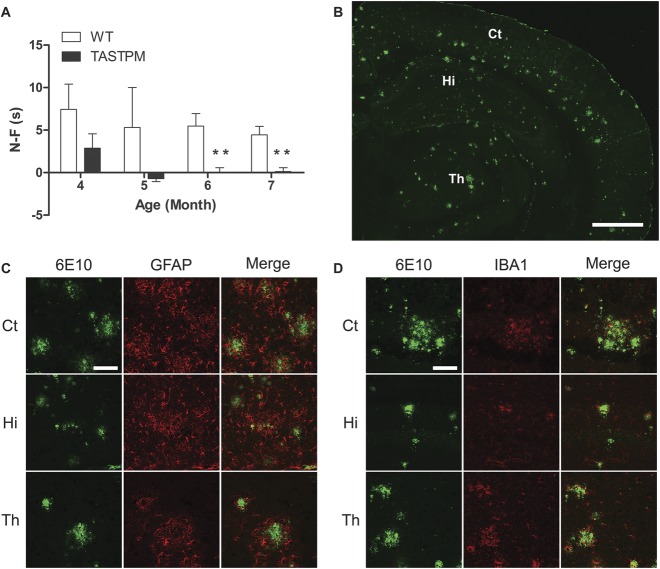
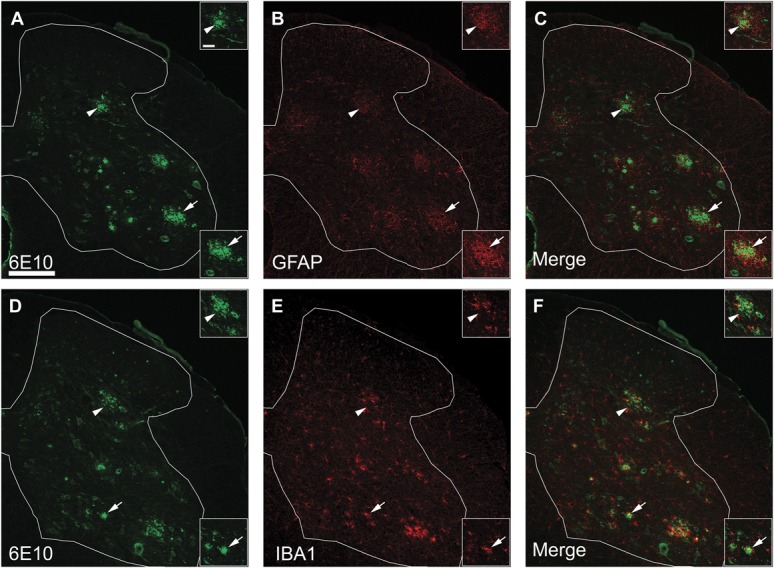
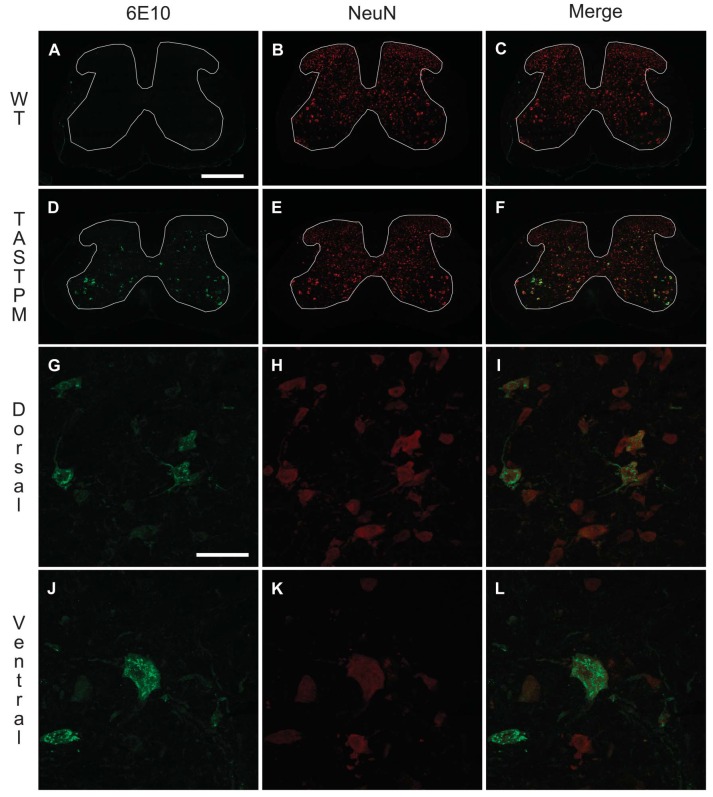
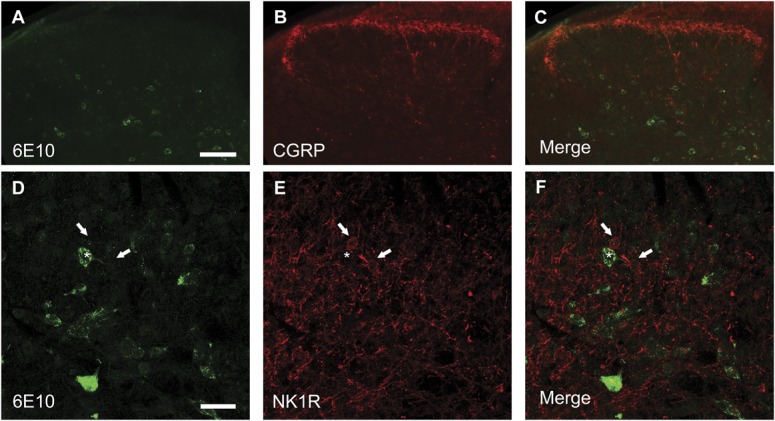
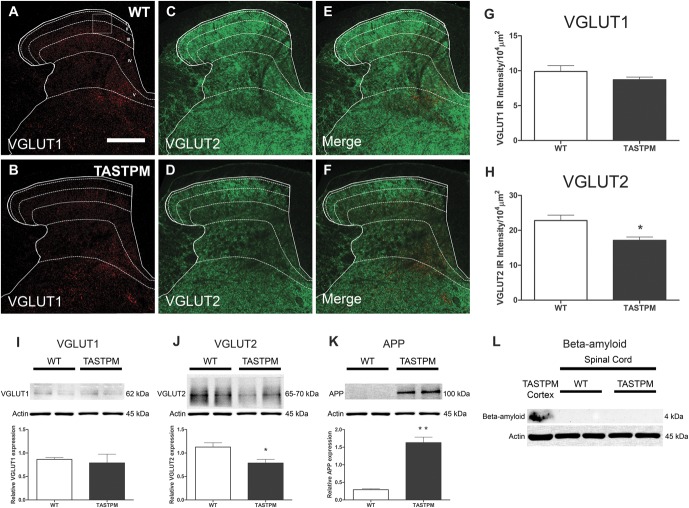
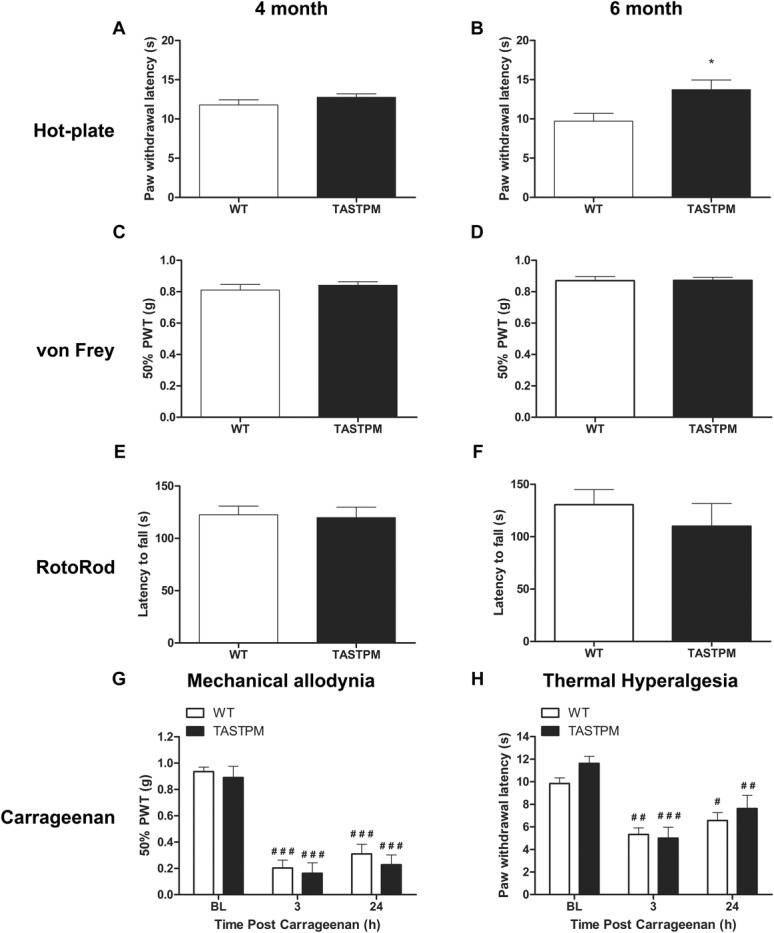
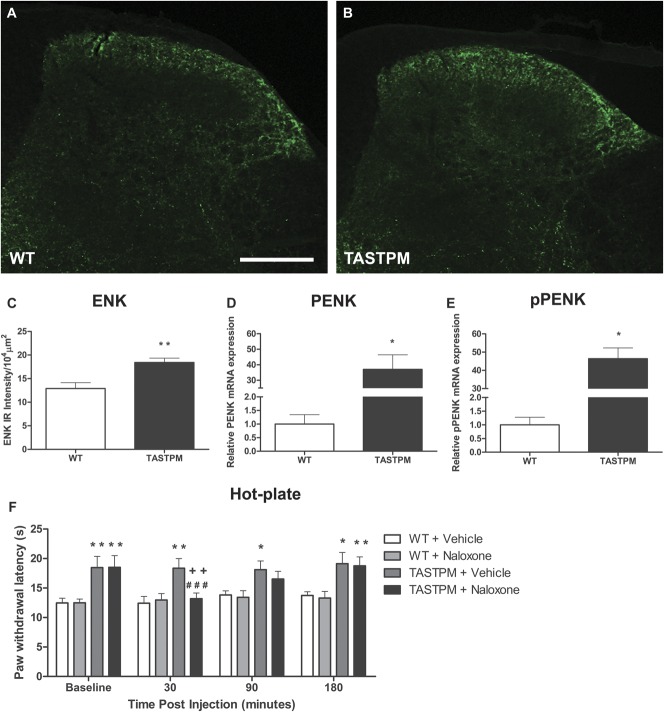
Individuals with Alzheimer's disease (AD) are in susceptible patient groups in which pain is an important clinical issue that is often underdiagnosed. However, it is unclear whether decreased pain complaints in patients with AD result from elevated pain tolerance or an impaired ability to communicate sensations. Here, we explored if AD-related pathology is present in key regions of the pain pathway and assessed whether nociceptive thresholds to acute noxious stimulation are altered in the double-mutant APPswe × PS1.M146V (TASTPM) transgenic mouse model of AD. TASTPM mice exhibited an age-dependant cognitive deficit at the age of 6 months, but not at 4 months, a deficit that was accompanied by amyloid plaques in the cortex, hippocampus, and thalamus. In the spinal cord, β-amyloid (APP/Aβ) immunoreactivity was observed in dorsal and ventral horn neurons, and the expression of vesicular glutamate transporter 2 (VGLUT2) was significantly reduced, while the expression of the inhibitory peptides enkephalins was increased in TASTPM dorsal horn, consistent with an increased inhibitory tone. TASTPM mice displayed reduced sensitivity to acute noxious heat, which was reversed by naloxone, an opioid antagonist. This study suggests that increased inhibition and decreased excitation in the spinal cord may be responsible for the reduced thermal sensitivity associated with AD-related pathology.
Conflict of interest statement
Sponsorships or competing interests that may be relevant to content are disclosed at the end of this article.
Figures
References
-
- Ballard CG, Gauthier S, Cummings JL, Brodaty H, Grossberg GT, Robert P, Lyketsos CG. Management of agitation and aggression associated with Alzheimer disease. Nat Rev Neurol 2009;5:245–55. - PubMed
-
- Benedetti F, Arduino C, Vighetti S, Asteggiano G, Tarenzi L, Rainero I. Pain reactivity in Alzheimer patients with different degrees of cognitive impairment and brain electrical activity deterioration. PAIN 2004;111:22–9. - PubMed
-
- Brooks SP, Dunnett SB. Tests to assess motor phenotype in mice: a user's guide. Nat Rev Neurosci 2009;10:519–29. - PubMed
-
- Brumovsky P, Watanabe M, Hokfelt T. Expression of the vesicular glutamate transporters-1 and -2 in adult mouse dorsal root ganglia and spinal cord and their regulation by nerve injury. Neuroscience 2007;147:469–90. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
