

Root cap-dependent gravitropic U-turn of maize root requires light-induced auxin biosynthesis via the YUC pathway in the root apex
- PMID: 27307546
- PMCID: PMC4973731
- DOI: 10.1093/jxb/erw232
Root cap-dependent gravitropic U-turn of maize root requires light-induced auxin biosynthesis via the YUC pathway in the root apex
Abstract
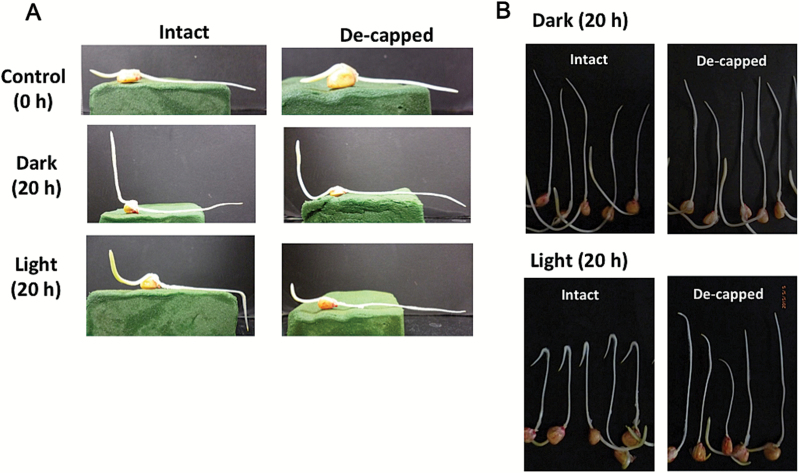
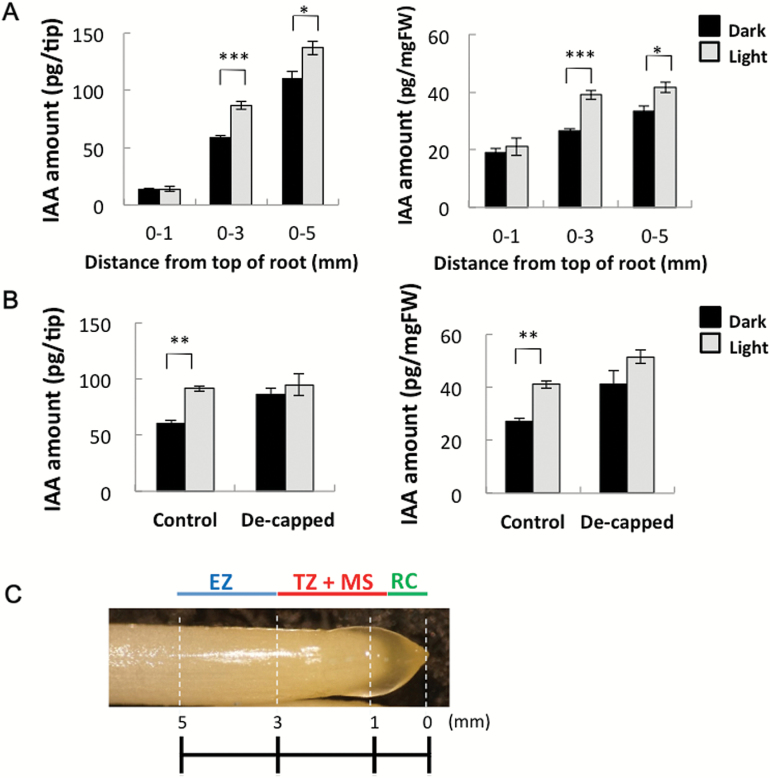
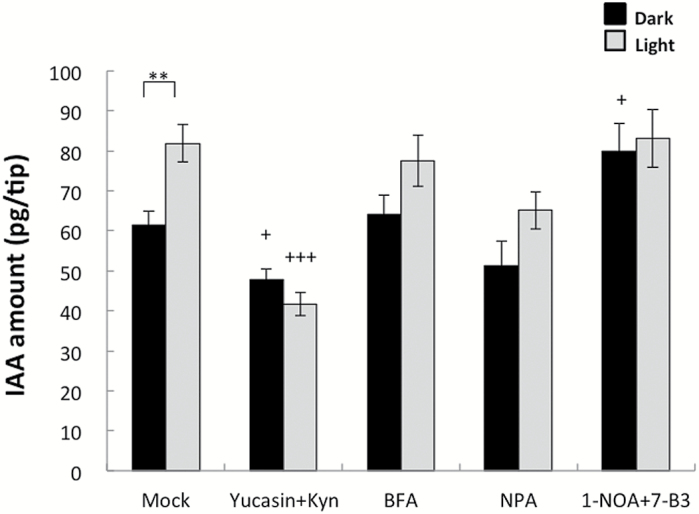
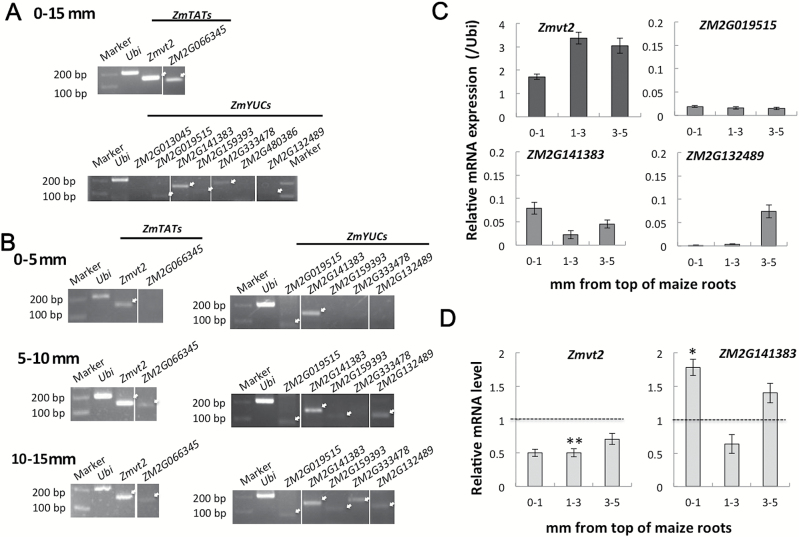
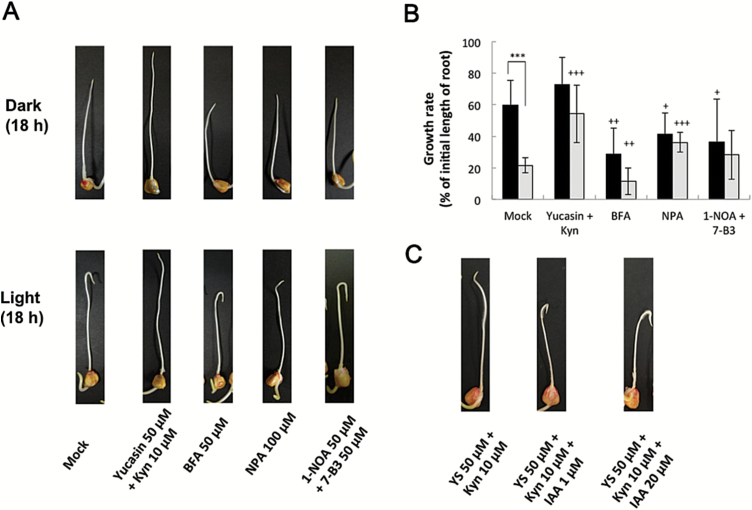
Gravitropism refers to the growth or movement of plants that is influenced by gravity. Roots exhibit positive gravitropism, and the root cap is thought to be the gravity-sensing site. In some plants, the root cap requires light irradiation for positive gravitropic responses. However, the mechanisms regulating this phenomenon are unknown. We herein report that maize roots exposed to white light continuously for ≥1-2h show increased indole-3-acetic acid (IAA) levels in the root tips, especially in the transition zone (1-3mm from the tip). Treatment with IAA biosynthesis inhibitors yucasin and l-kynurenine prevented any increases in IAA content and root curvature under light conditions. Analyses of the incorporation of a stable isotope label from tryptophan into IAA revealed that some of the IAA in roots was synthesized in the root apex. Furthermore, Zmvt2 and Zmyuc gene transcripts were detected in the root apex. One of the Zmyuc genes (ZM2G141383) was up-regulated by light irradiation in the 0-1mm tip region. Our findings suggest that IAA accumulation in the transition zone is due to light-induced activation of Zmyuc gene expression in the 0-1mm root apex region. Light-induced changes in IAA levels and distributions mediate the maize root gravitropic U-turn.
Keywords: Indole-3-acetic acid (IAA) biosynthesis; Zmvt2; Zmyuc.; light dependent; maize; root cap; root gravitropism; yucasin.
© The Author 2016. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures


References
-
- Berleth T, Sachs T. 2001. Plant morphogenesis: long-distance coordination and local patterning. Current Opinion in Plant Biology 4, 57–62. - PubMed
-
- Brunoud G, Wells DM, Oliva M, et al. 2012. A novel sensor to map auxin response and distribution at high spatio-temporal resolution. Nature 482, 103–106. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
