

Molecular architecture of the human sperm IZUMO1 and egg JUNO fertilization complex
- PMID: 27309818
- PMCID: PMC5319863
- DOI: 10.1038/nature18595
Molecular architecture of the human sperm IZUMO1 and egg JUNO fertilization complex
Abstract
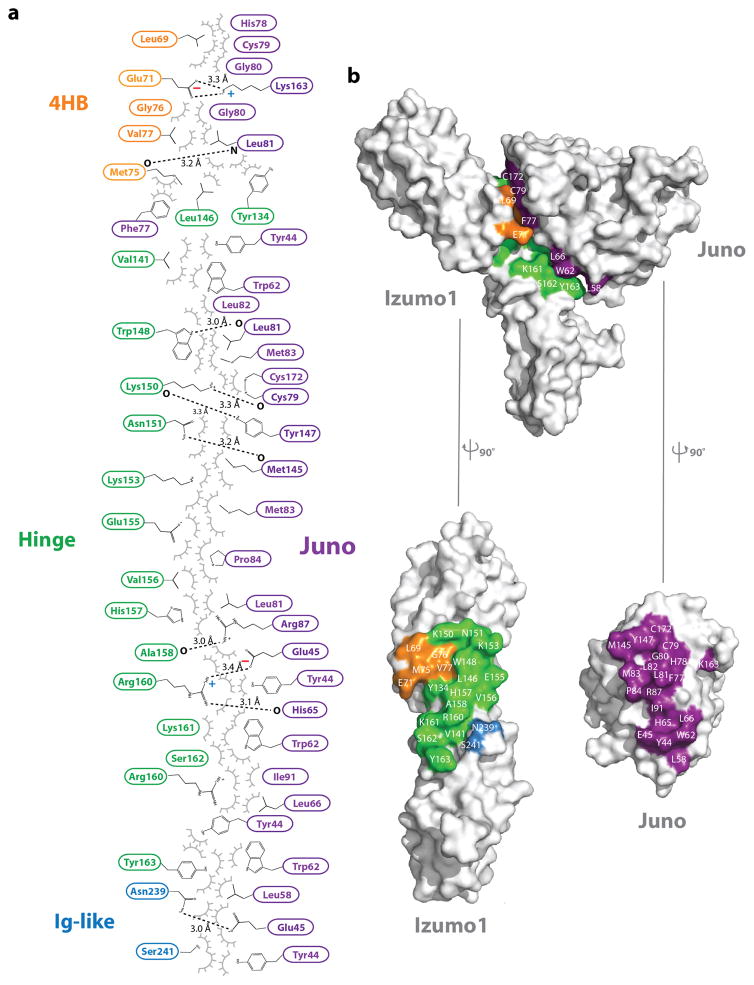
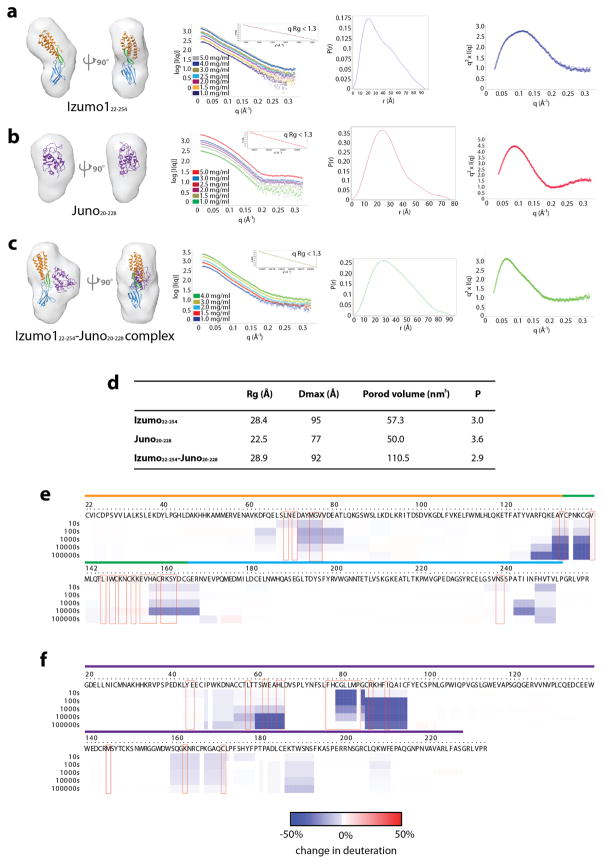
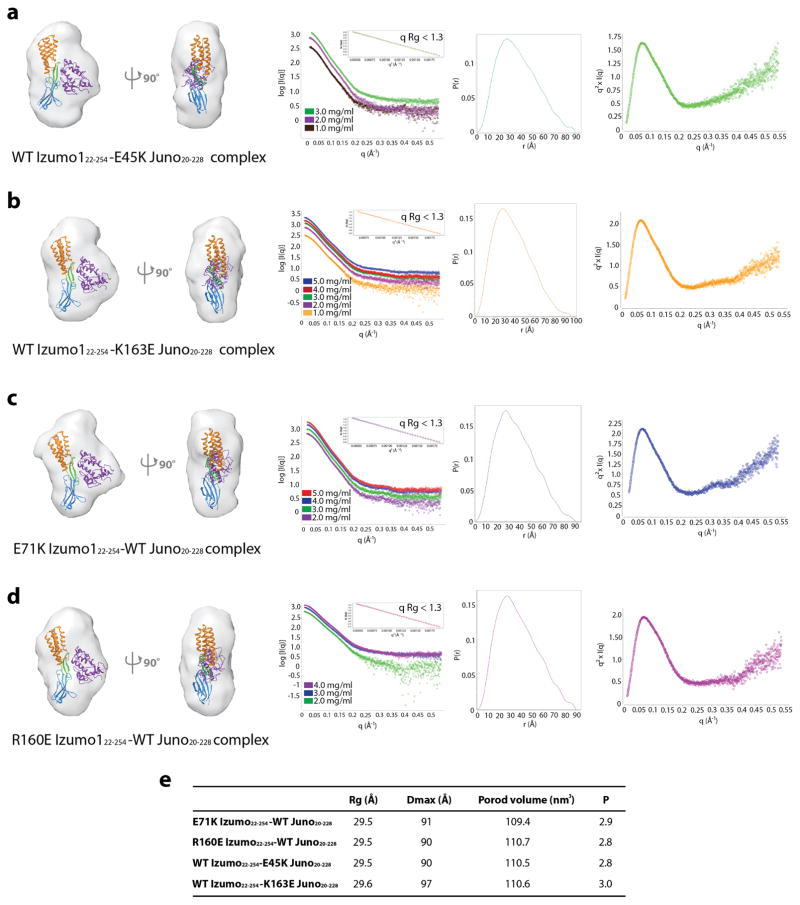
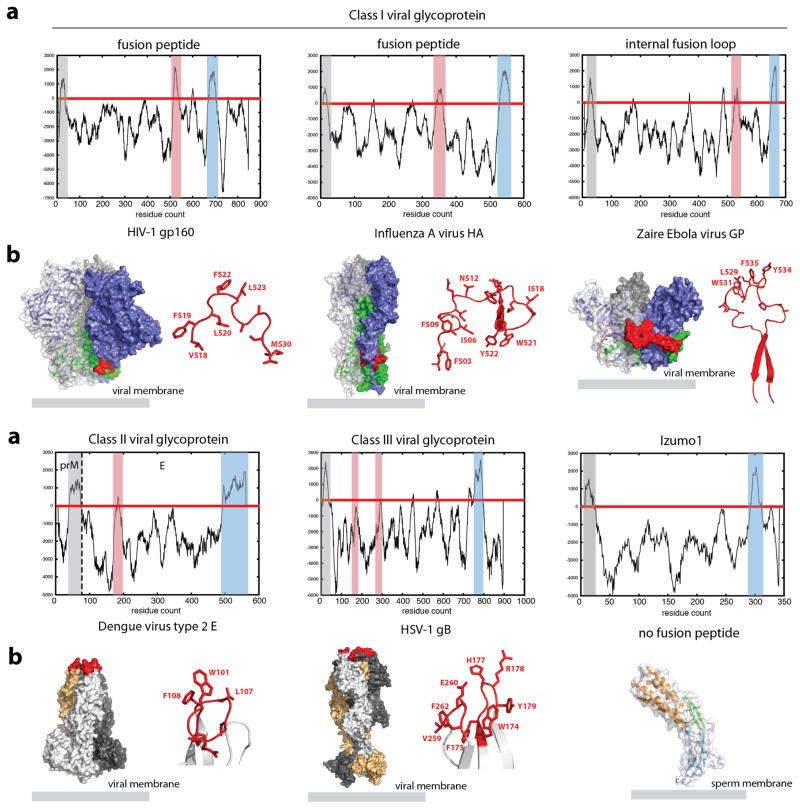
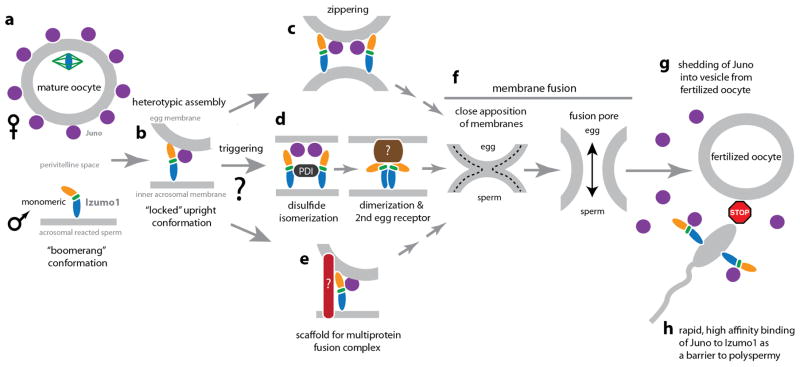
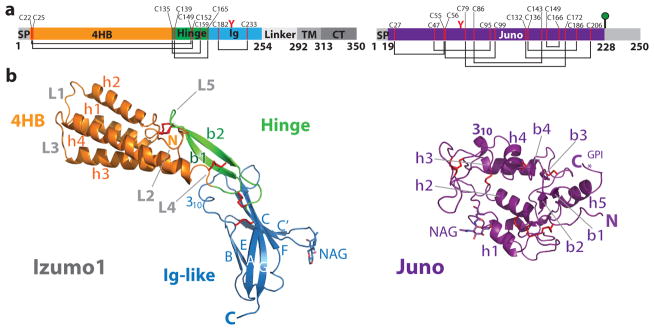
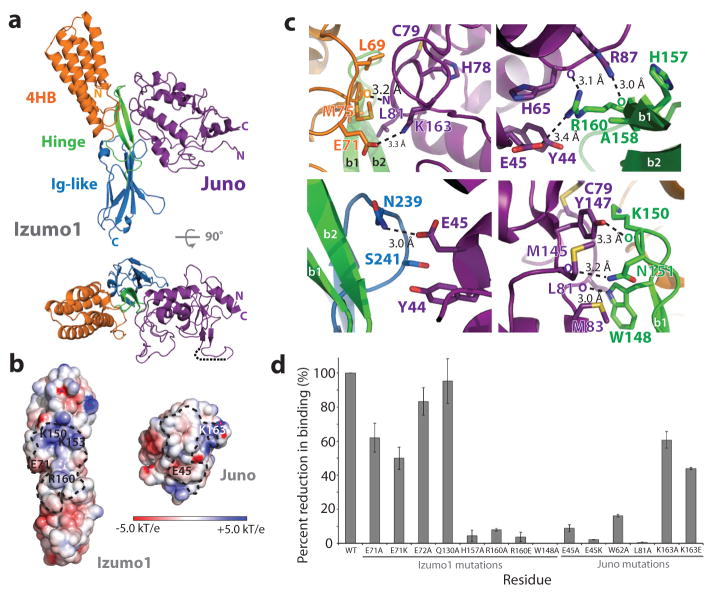
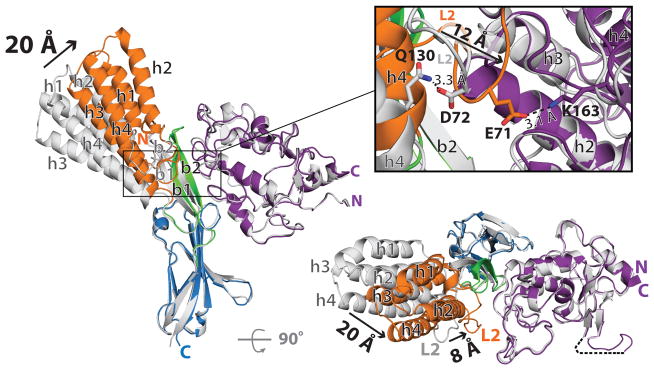
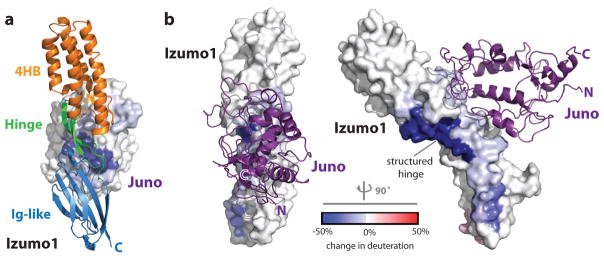
Fertilization is an essential biological process in sexual reproduction and comprises a series of molecular interactions between the sperm and egg. The fusion of the haploid spermatozoon and oocyte is the culminating event in mammalian fertilization, enabling the creation of a new, genetically distinct diploid organism. The merger of two gametes is achieved through a two-step mechanism in which the sperm protein IZUMO1 on the equatorial segment of the acrosome-reacted sperm recognizes its receptor, JUNO, on the egg surface. This recognition is followed by the fusion of the two plasma membranes. IZUMO1 and JUNO proteins are indispensable for fertilization, as constitutive knockdown of either protein results in mice that are healthy but infertile. Despite their central importance in reproductive medicine, the molecular architectures of these proteins and the details of their functional roles in fertilization are not known. Here we present the crystal structures of human IZUMO1 and JUNO in unbound and bound conformations. The human IZUMO1 structure exhibits a distinct boomerang shape and provides structural insights into the IZUMO family of proteins. Human IZUMO1 forms a high-affinity complex with JUNO and undergoes a major conformational change within its N-terminal domain upon binding to the egg-surface receptor. Our results provide insights into the molecular basis of sperm-egg recognition, cross-species fertilization, and the barrier to polyspermy, thereby promising benefits for the rational development of non-hormonal contraceptives and fertility treatments for humans and other mammals.
Figures
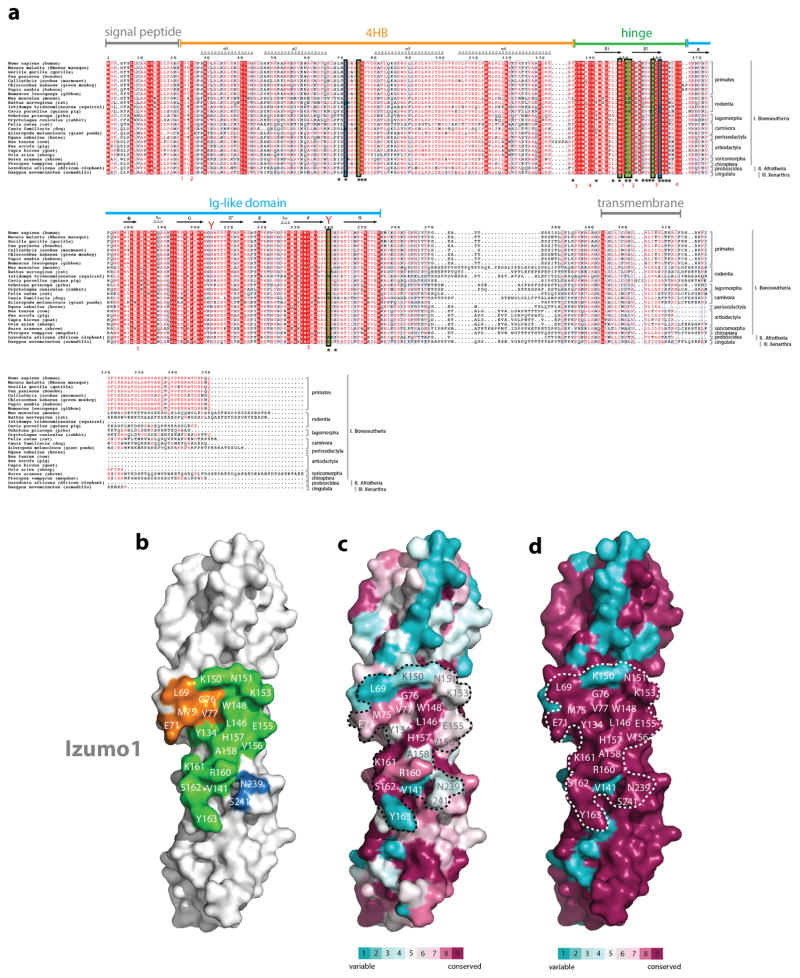
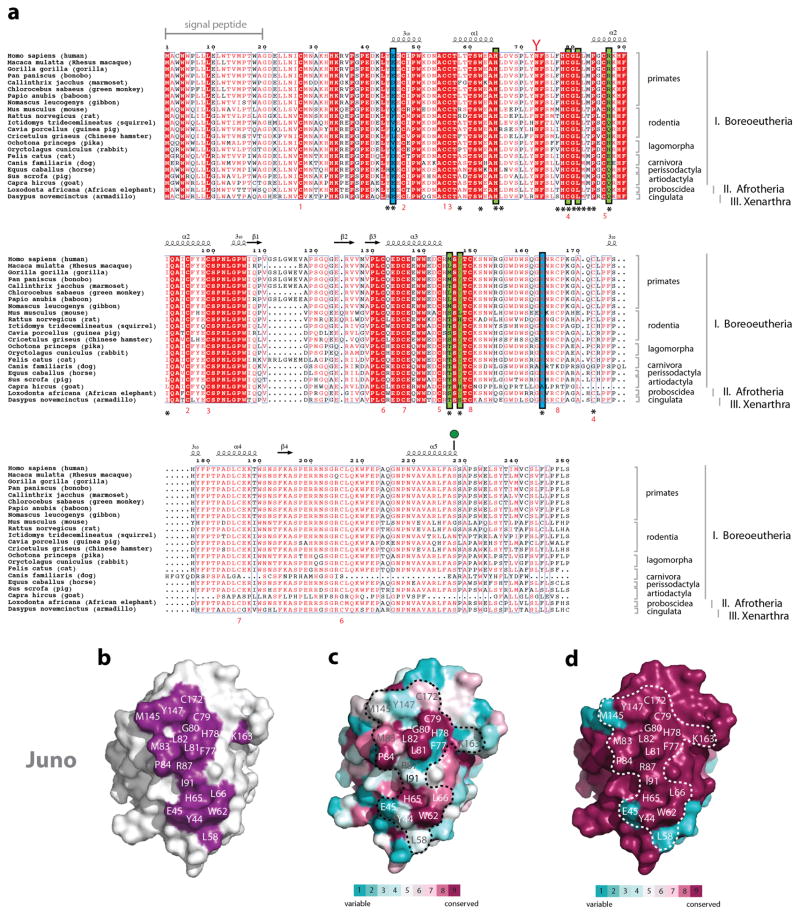
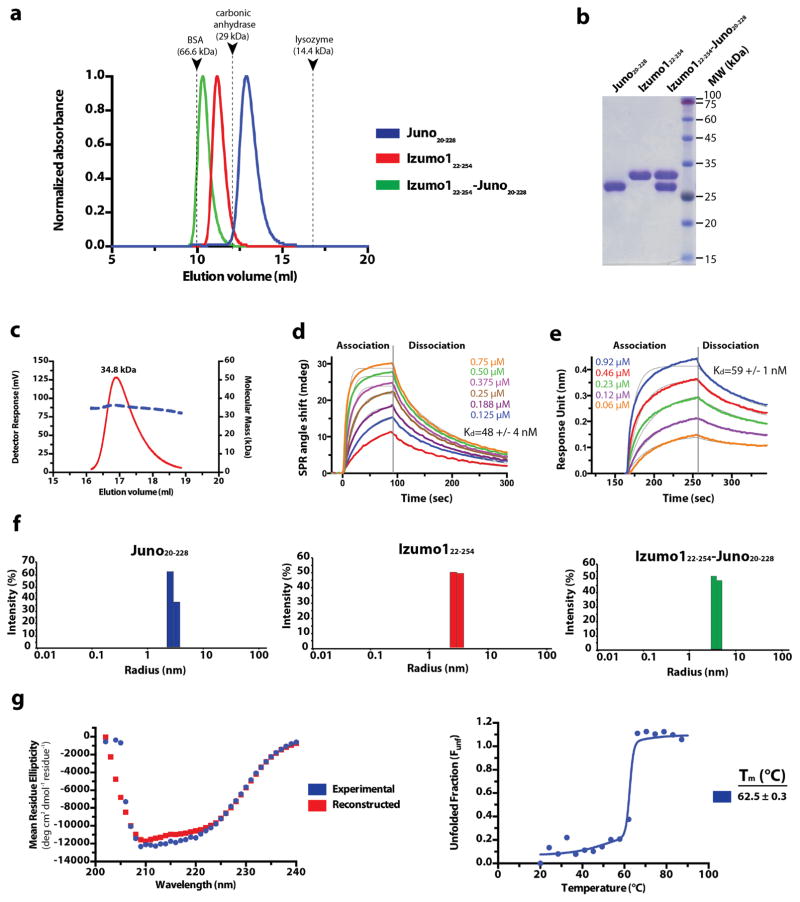
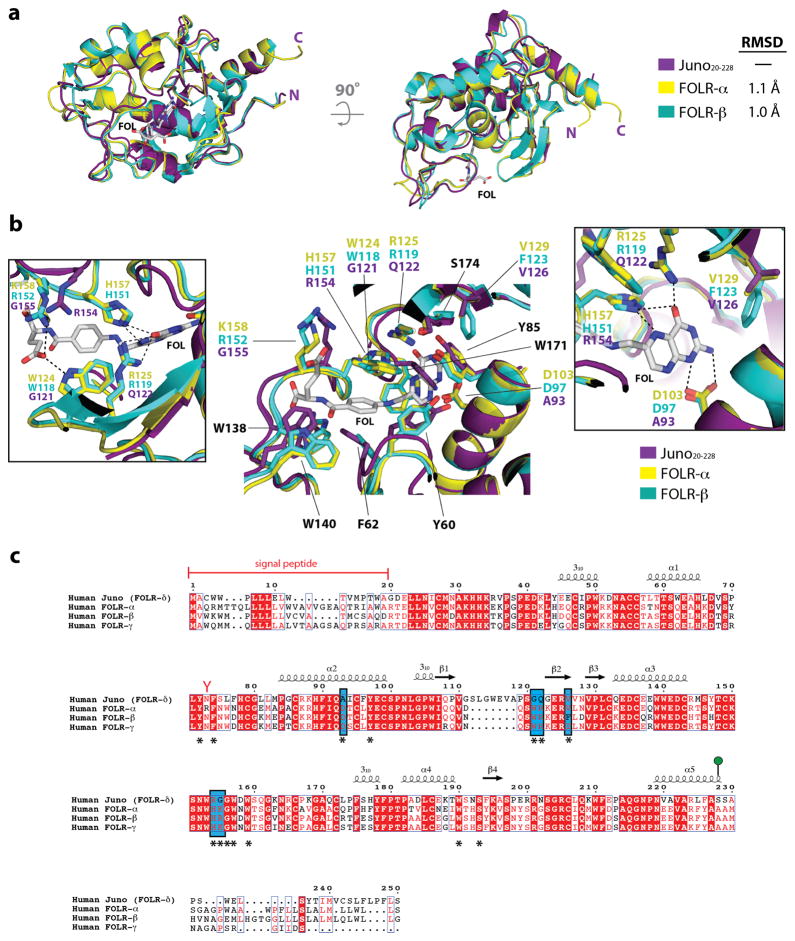
Comment in
-
Structural biology: When sperm meets egg.Nature. 2016 Jun 23;534(7608):484-5. doi: 10.1038/nature18448. Epub 2016 Jun 15. Nature. 2016. PMID: 27309810 No abstract available.
References
-
- Wassarman PM, Jovine L, Litscher ES. A profile of fertilization in mammals. Nat Cell Biol. 2001;3:E59–E64. - PubMed
-
- Evans JP. Sperm-Egg Interaction. Annu Rev Physiol. 2012;74:477–502. - PubMed
-
- Sutovsky P. Sperm–egg adhesion and fusion in mammals. Expert Rev Mol Med. 2009;11 - PubMed
-
- Inoue N, Ikawa M, Isotani A, Okabe M. The immunoglobulin superfamily protein Izumo is required for sperm to fuse with eggs. Nature. 2005;434:234–238. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
