

Filamentation of Metabolic Enzymes in Saccharomyces cerevisiae
- PMID: 27312010
- PMCID: PMC4920916
- DOI: 10.1016/j.jgg.2016.03.008
Filamentation of Metabolic Enzymes in Saccharomyces cerevisiae
Abstract
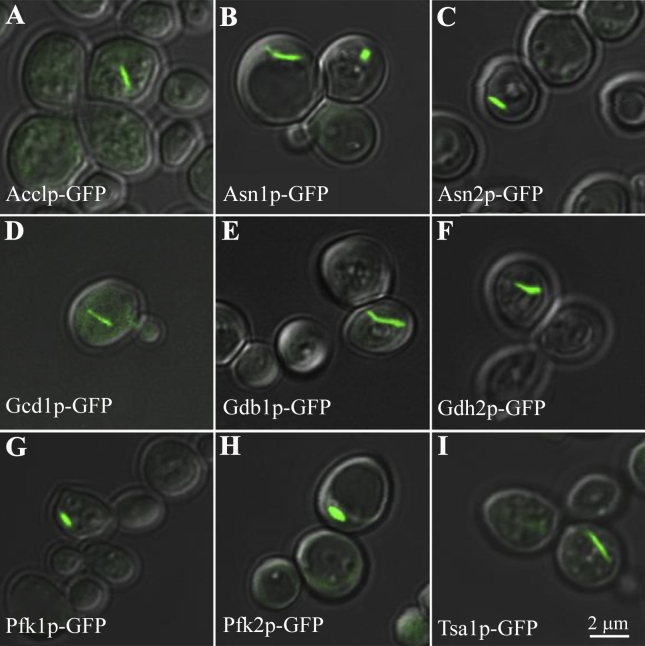
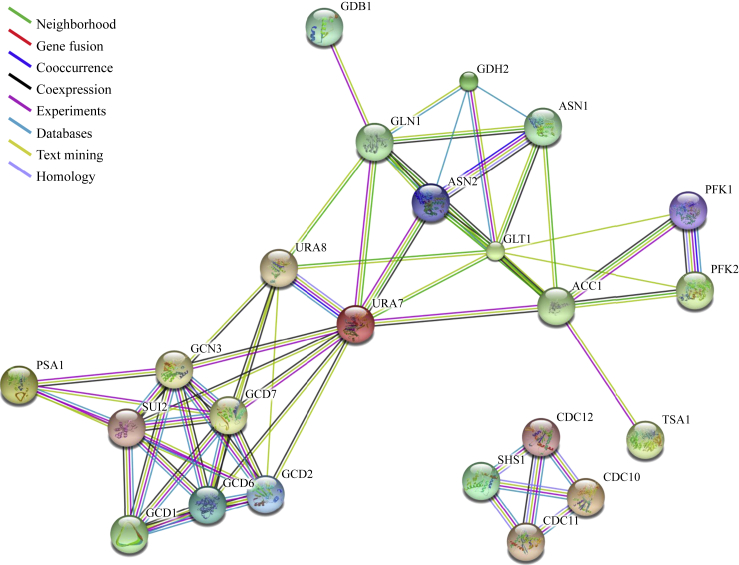
Compartmentation via filamentation has recently emerged as a novel mechanism for metabolic regulation. In order to identify filament-forming metabolic enzymes systematically, we performed a genome-wide screening of all strains available from an open reading frame-GFP collection in Saccharomyces cerevisiae. We discovered nine novel filament-forming proteins and also confirmed those identified previously. From the 4159 strains, we found 23 proteins, mostly metabolic enzymes, which are capable of forming filaments in vivo. In silico protein-protein interaction analysis suggests that these filament-forming proteins can be clustered into several groups, including translational initiation machinery and glucose and nitrogen metabolic pathways. Using glutamine-utilising enzymes as examples, we found that the culture conditions affect the occurrence and length of the metabolic filaments. Furthermore, we found that two CTP synthases (Ura7p and Ura8p) and two asparagine synthetases (Asn1p and Asn2p) form filaments both in the cytoplasm and in the nucleus. Live imaging analyses suggest that metabolic filaments undergo sub-diffusion. Taken together, our genome-wide screening identifies additional filament-forming proteins in S. cerevisiae and suggests that filamentation of metabolic enzymes is more general than currently appreciated.
Keywords: CTP synthase; Cytoophidium; Glutamine; Glycolysis; Intracellular compartmentation; Metabolic enzyme; Saccharomyces cerevisiae.
Copyright © 2016 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures



References
-
- An S., Kumar R., Sheets E.D., Benkovic S.J. Reversible compartmentalization of de novo purine biosynthetic complexes in living cells. Science. 2008;320:103–106. - PubMed
-
- Aughey G.N., Tastan Ö.Y., Liu J.L. Cellular serpents and dreaming spires: new frontiers in arginine and pyrimidine biology. J. Genet. Genomics. 2014;41:561–565. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
