

A novel TP53 pathway influences the HGS-mediated exosome formation in colorectal cancer
- PMID: 27312428
- PMCID: PMC4911576
- DOI: 10.1038/srep28083
A novel TP53 pathway influences the HGS-mediated exosome formation in colorectal cancer
Abstract
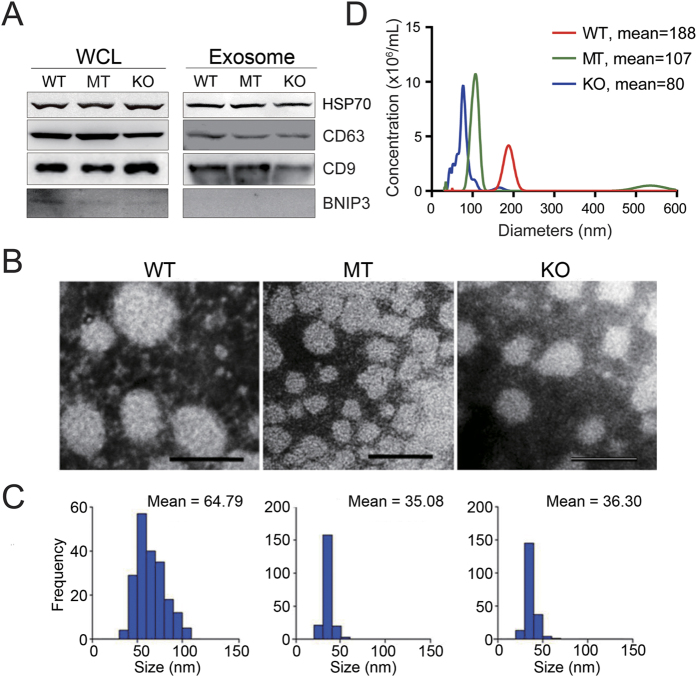
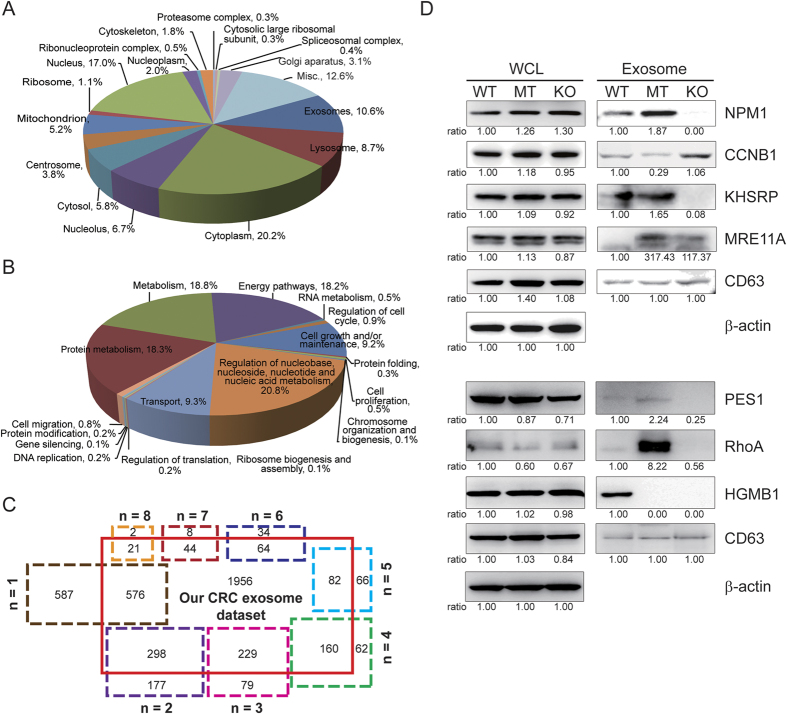
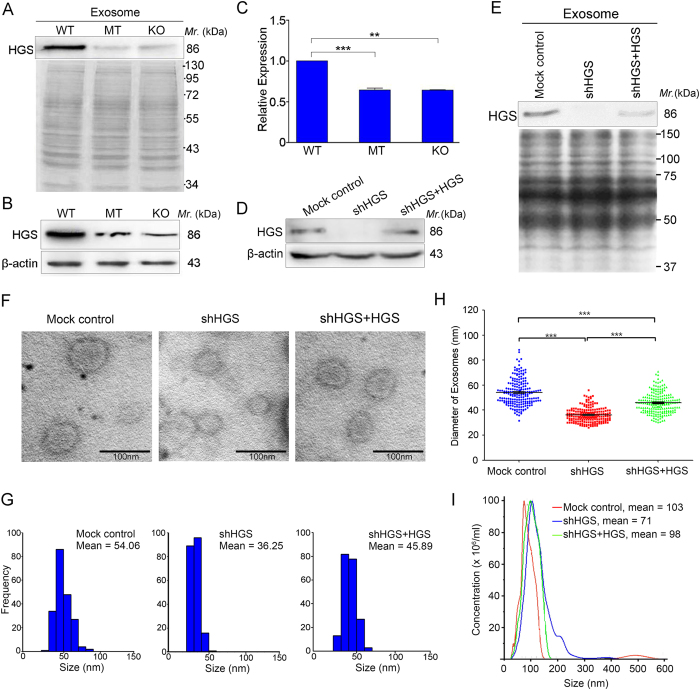
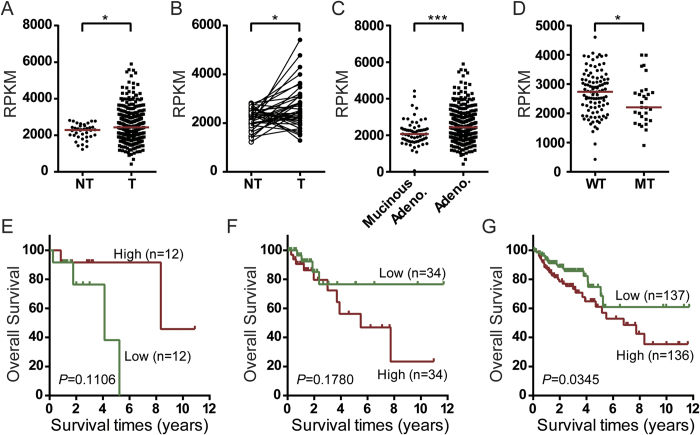
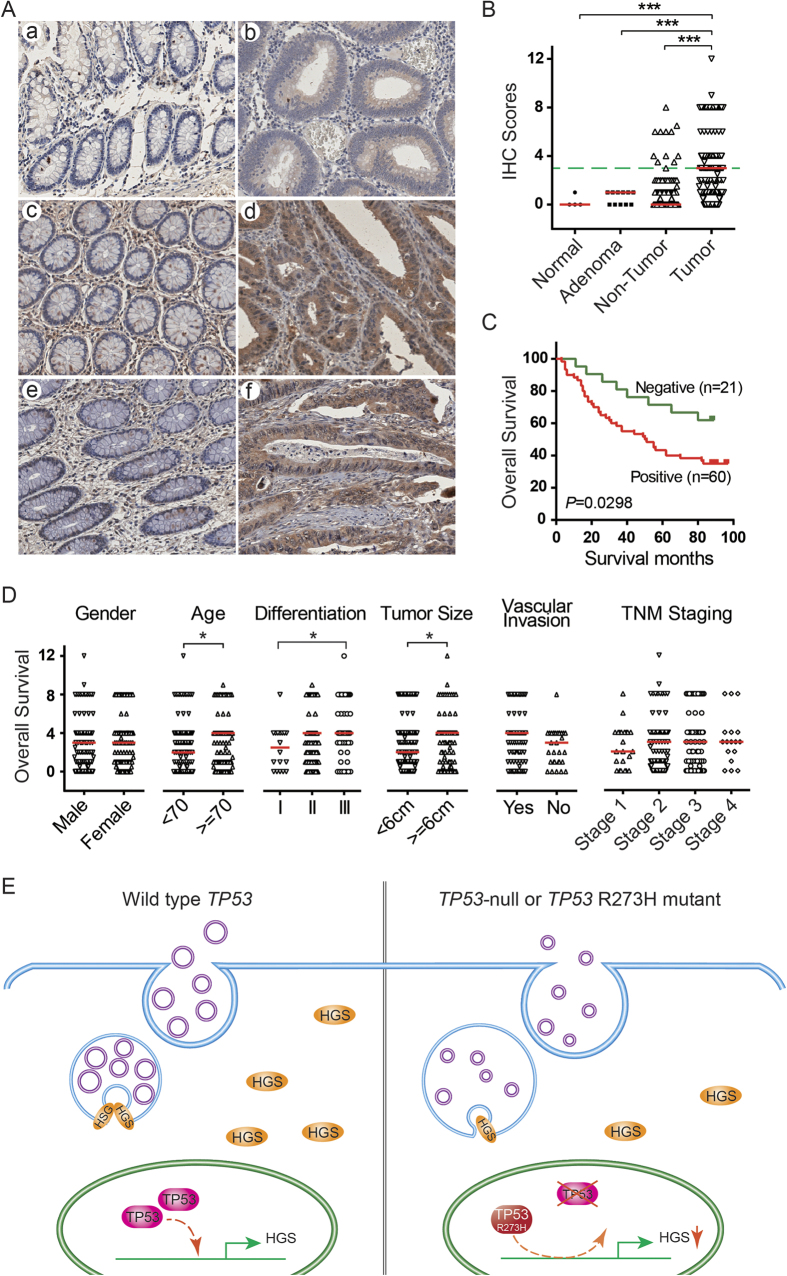
Tumor-derived exosomes are important for cell-cell communication. However, the role of TP53 in the control of exosome production in colorectal cancer (CRC) is controversial and unclear. The features of exosomes secreted from HCT116 TP53-wild type (WT), TP53-knockout (KO) and constructed TP53 (R273H)-mutant (MT) cells were assessed. The exosomes from the MT and KO cells exhibited significantly reduced sizes compared with the WT cells. A comprehensive proteomic analysis of exosomal proteins was performed using the isobaric tag for relative and absolute quantitation (iTRAQ)-2D-LC-MS/MS strategy. A total of 3437 protein groups with ≥2 matched peptides were identified. Specifically, hepatocyte growth factor-regulated tyrosine kinase substrate (HGS) was consistently down-regulated in the exosomes from the MT and KO cells. Functional studies demonstrated that low HGS levels were responsible for the decreased exosome size. TP53 regulated HGS expression and thus HGS-dependent exosome formation. Furthermore, the HGS expression was gradually increased concomitant with CRC carcinogenesis and was an independent poor prognostic factor. In conclusion, a novel HGS-dependent TP53 mechanism in exosome formation was identified in CRC. HGS may serve as a novel prognostic biomarker and a candidate target for therapeutic interventions.
Figures
References
-
- Jemal A. et al. Global cancer statistics. CA Cancer J. Clin. 61, 69–90 (2011). - PubMed
-
- Fearon E. R. & Vogelstein B. A genetic model for colorectal tumorigenesis. Cell 61, 759–767 (1990). - PubMed
-
- Brosh R. & Rotter V. When mutants gain new powers: news from the mutant p53 field. Nature reviews. Cancer 9, 701–713 (2009). - PubMed
-
- Naccarati A. et al. Mutations and polymorphisms in TP53 gene–an overview on the role in colorectal cancer. Mutagenesis 27, 211–218 (2012). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
