

Translational control by 5'-untranslated regions of eukaryotic mRNAs
- PMID: 27313038
- PMCID: PMC7422601
- DOI: 10.1126/science.aad9868
Translational control by 5'-untranslated regions of eukaryotic mRNAs
Abstract
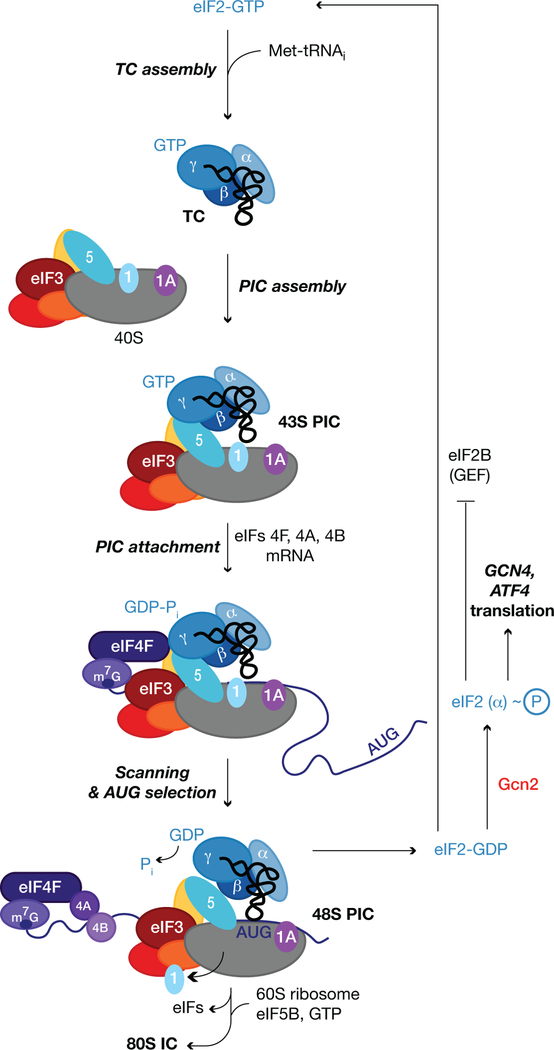
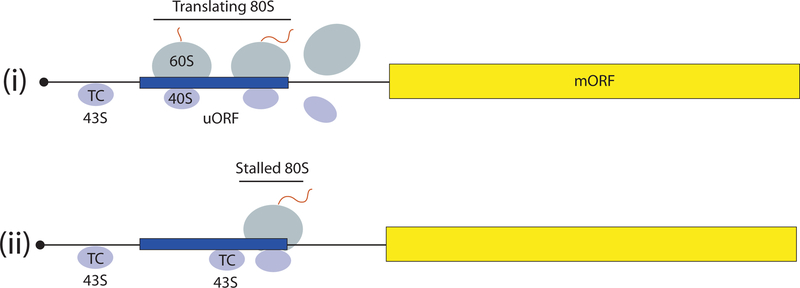
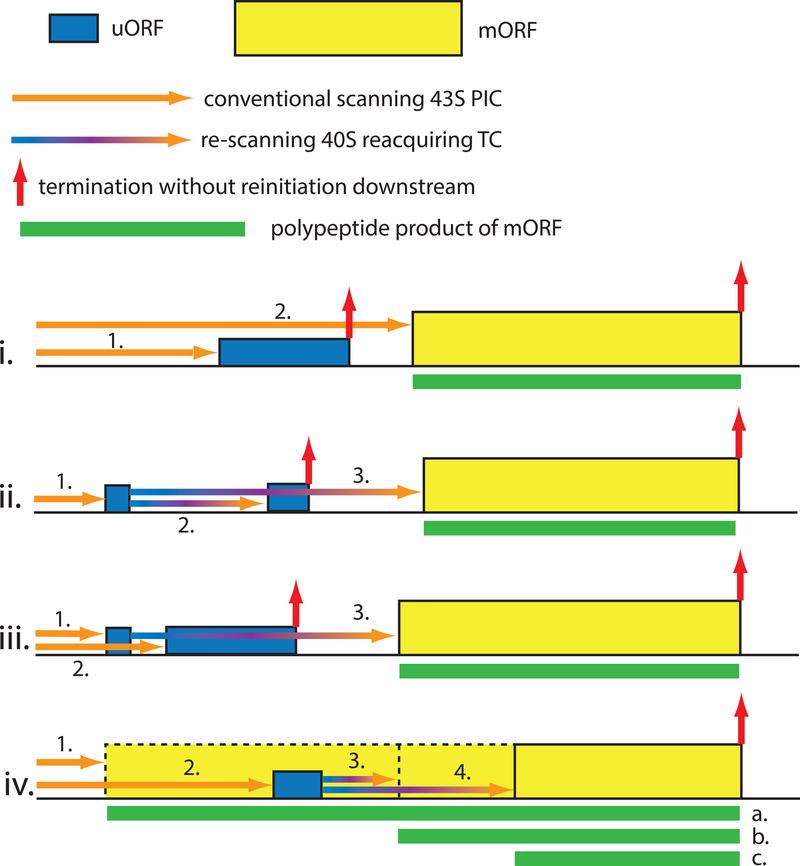
The eukaryotic 5' untranslated region (UTR) is critical for ribosome recruitment to the messenger RNA (mRNA) and start codon choice and plays a major role in the control of translation efficiency and shaping the cellular proteome. The ribosomal initiation complex is assembled on the mRNA via a cap-dependent or cap-independent mechanism. We describe various mechanisms controlling ribosome scanning and initiation codon selection by 5' upstream open reading frames, translation initiation factors, and primary and secondary structures of the 5'UTR, including particular sequence motifs. We also discuss translational control via phosphorylation of eukaryotic initiation factor 2, which is implicated in learning and memory, neurodegenerative diseases, and cancer.
Copyright © 2016, American Association for the Advancement of Science.
Figures
References
-
- Kozak M, Point mutations define a sequence flanking the AUG initiator codon that modulates translation by eukaryotic ribosomes. Cell 44, 283–292 (1986). - PubMed
-
- Starck SR et al., Leucine-tRNA initiates at CUG start codons for protein synthesis and presentation by MHC class I. Science 336, 1719–1723 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
