

Biosensor reveals multiple sources for mitochondrial NAD⁺
- PMID: 27313049
- PMCID: PMC6530784
- DOI: 10.1126/science.aad5168
Biosensor reveals multiple sources for mitochondrial NAD⁺
Abstract
Nicotinamide adenine dinucleotide (NAD(+)) is an essential substrate for sirtuins and poly(adenosine diphosphate-ribose) polymerases (PARPs), which are NAD(+)-consuming enzymes localized in the nucleus, cytosol, and mitochondria. Fluctuations in NAD(+) concentrations within these subcellular compartments are thought to regulate the activity of NAD(+)-consuming enzymes; however, the challenge in measuring compartmentalized NAD(+) in cells has precluded direct evidence for this type of regulation. We describe the development of a genetically encoded fluorescent biosensor for directly monitoring free NAD(+) concentrations in subcellular compartments. We found that the concentrations of free NAD(+) in the nucleus, cytoplasm, and mitochondria approximate the Michaelis constants for sirtuins and PARPs in their respective compartments. Systematic depletion of enzymes that catalyze the final step of NAD(+) biosynthesis revealed cell-specific mechanisms for maintaining mitochondrial NAD(+) concentrations.
Copyright © 2016, American Association for the Advancement of Science.
Figures
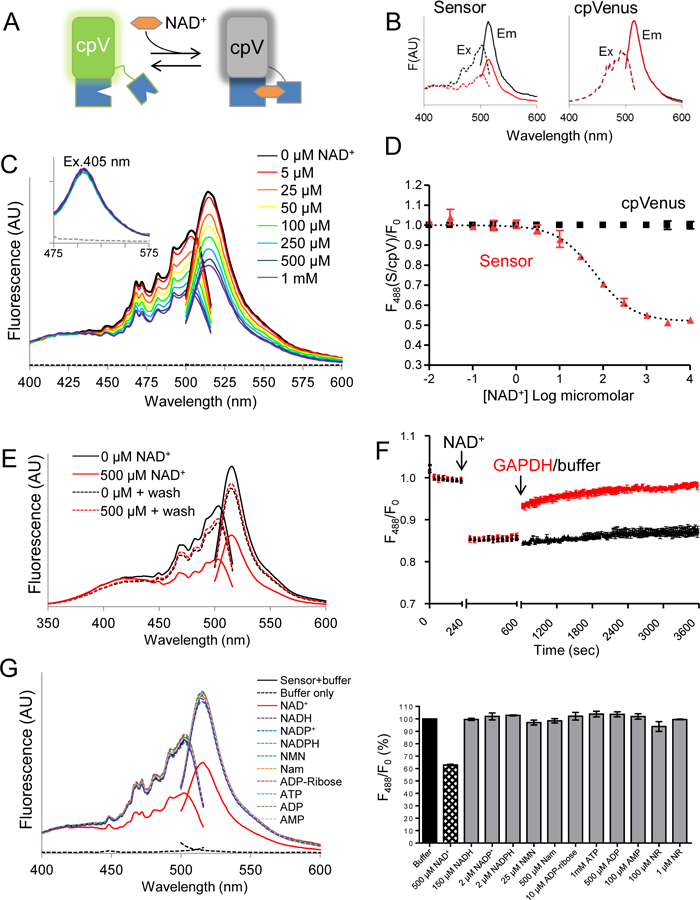
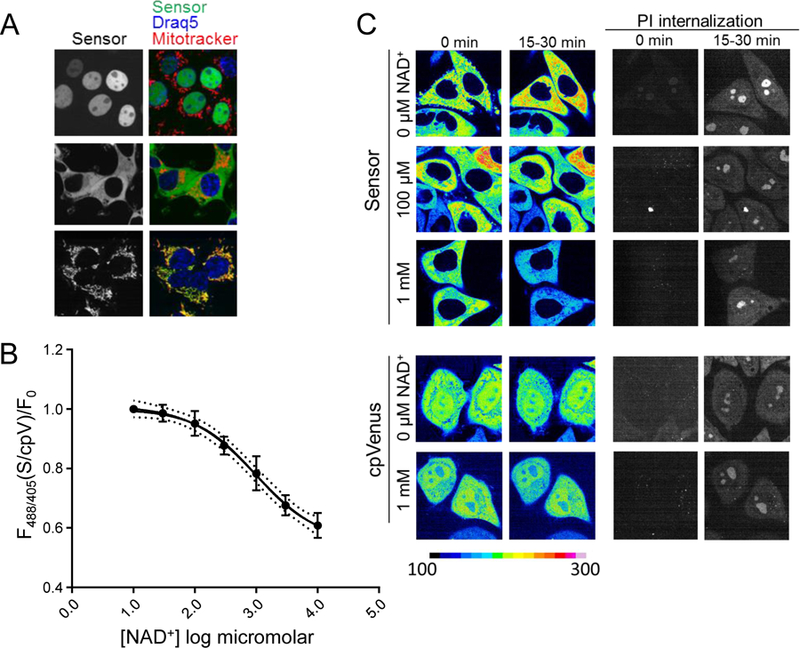

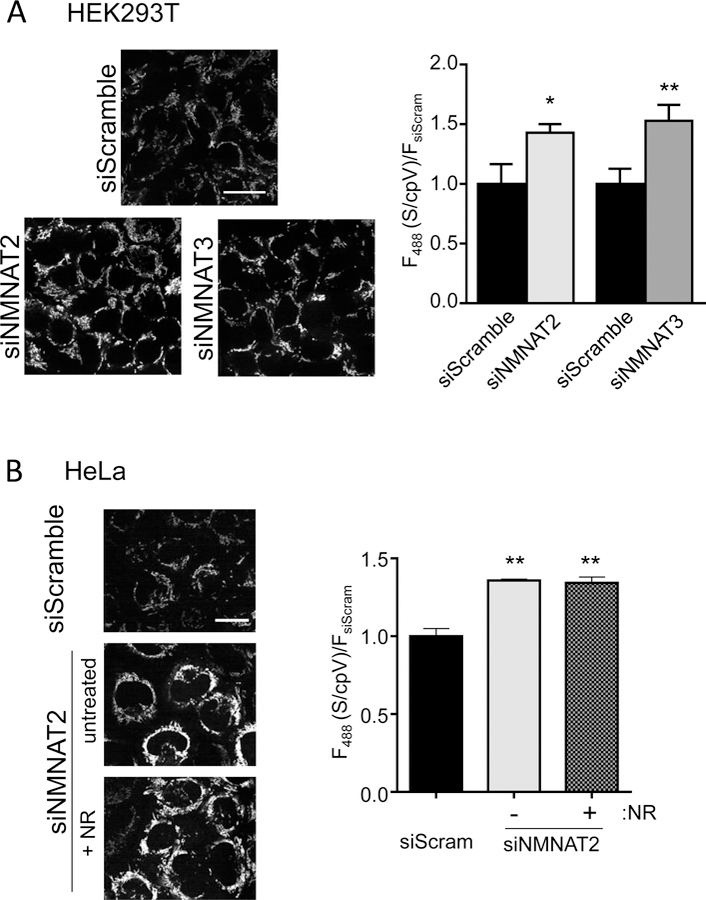
Comment in
-
CELL METABOLISM. The resurgence of NAD⁺.Science. 2016 Jun 17;352(6292):1396-7. doi: 10.1126/science.aag1718. Science. 2016. PMID: 27313027 No abstract available.
References
-
- Sauve AA, Celic I, Avalos J, Deng H, Boeke JD, et al. Chemistry of gene silencing: the mechanism of NAD+-dependent deacetylation reactions. Biochemistry 40, 15456–15463 (2001). - PubMed
-
- Revollo JR, Grimm AA, Imai S. The NAD biosynthesis pathway mediated by nicotinamide phosphoribosyltransferase regulates Sir2 activity in mammalian cells. J Biol Chem 279, 50754–50763 (2004). - PubMed
-
- Berger F, Lau C, Dahlmann M, Ziegler M. Subcellular compartmentation and differential catalytic properties of the three human nicotinamide mononucleotideadenylyltransferase isoforms. J Biol Chem 280, 36334–36341 (2005). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
