

VgrG C terminus confers the type VI effector transport specificity and is required for binding with PAAR and adaptor-effector complex
- PMID: 27313214
- PMCID: PMC4941472
- DOI: 10.1073/pnas.1600428113
VgrG C terminus confers the type VI effector transport specificity and is required for binding with PAAR and adaptor-effector complex
Abstract
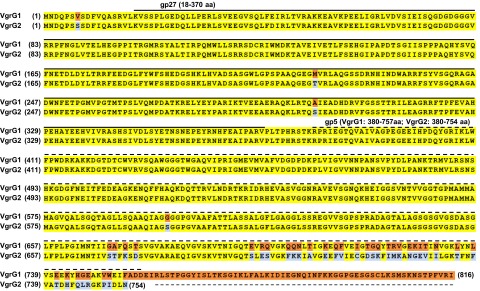
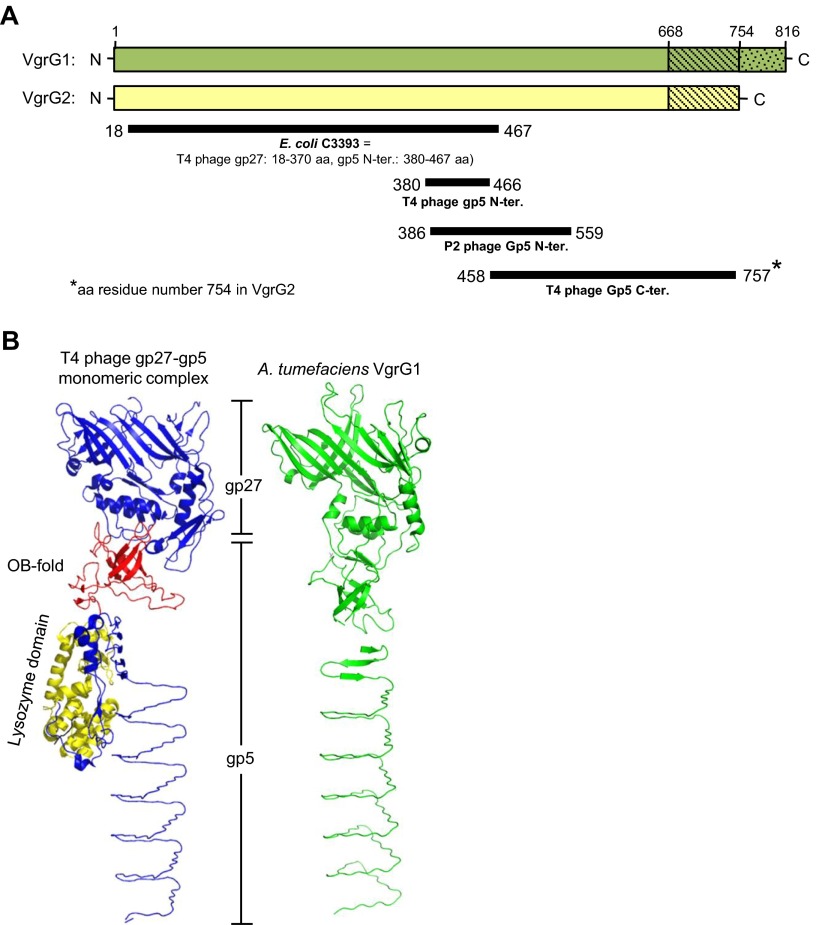
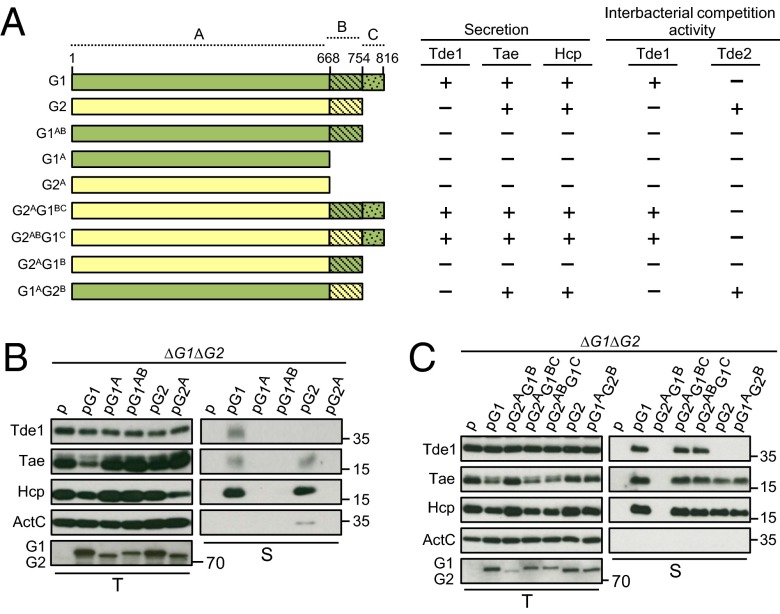
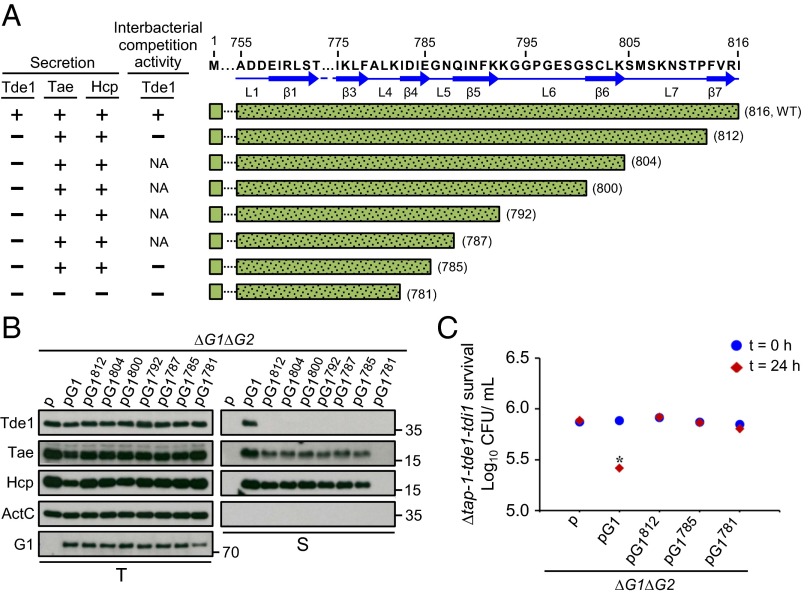
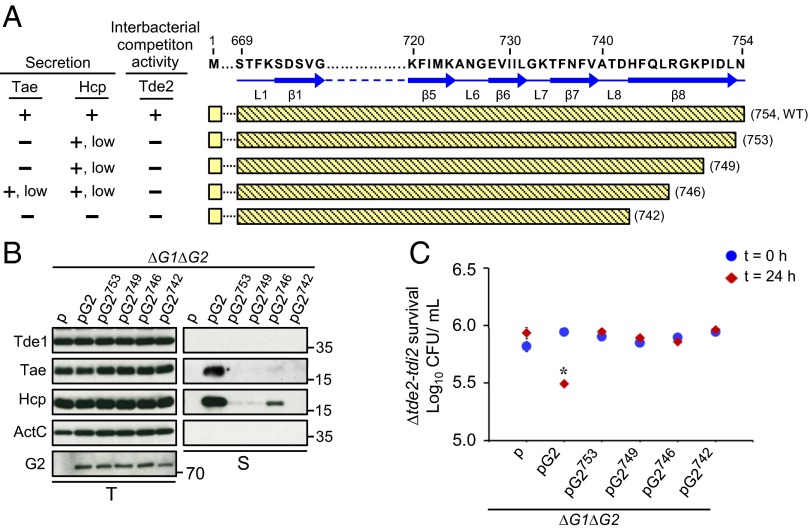
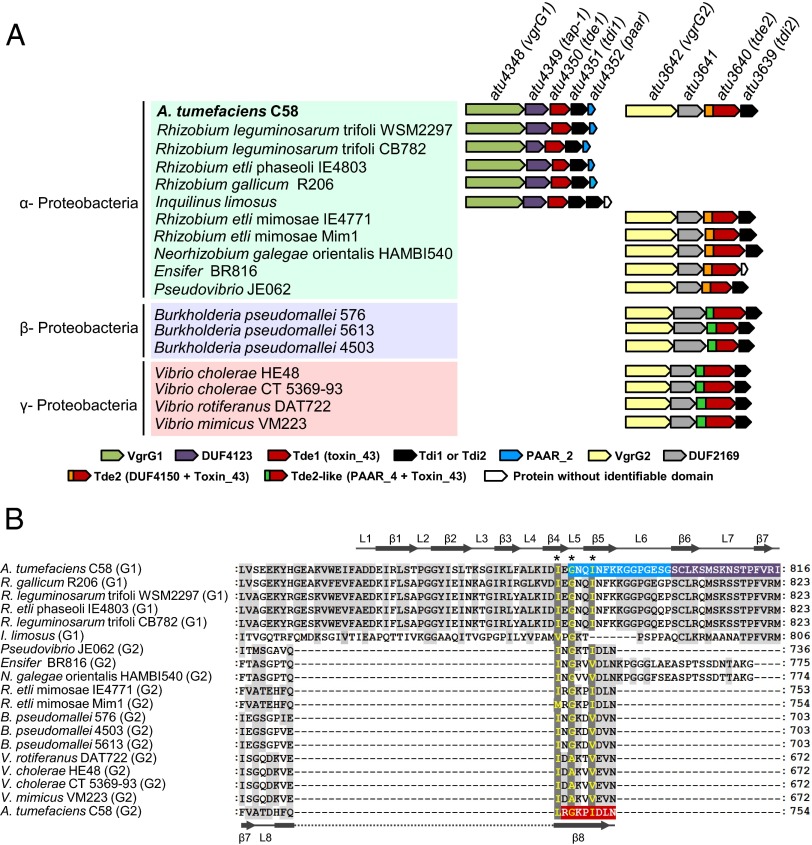
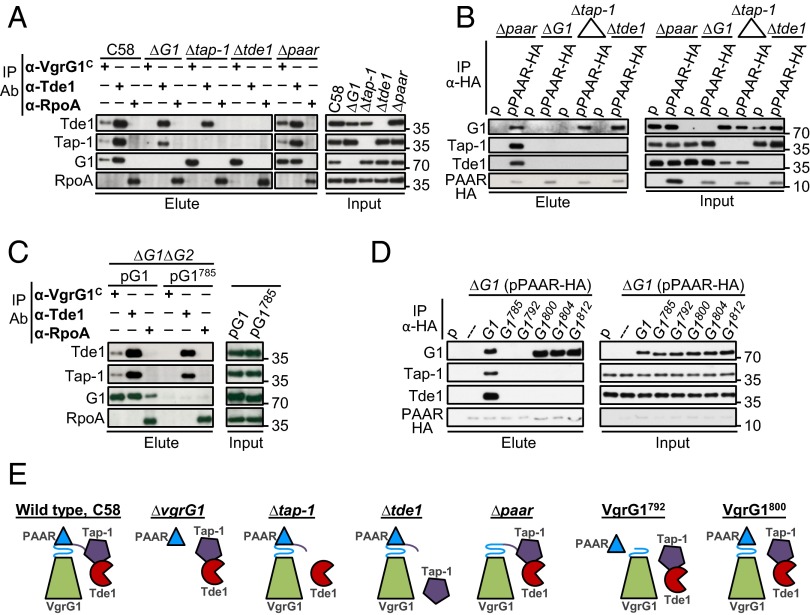
Type VI secretion system (T6SS) is a macromolecular machine used by many Gram-negative bacteria to inject effectors/toxins into eukaryotic hosts or prokaryotic competitors for survival and fitness. To date, our knowledge of the molecular determinants and mechanisms underlying the transport of these effectors remains limited. Here, we report that two T6SS encoded valine-glycine repeat protein G (VgrG) paralogs in Agrobacterium tumefaciens C58 specifically control the secretion and interbacterial competition activity of the type VI DNase toxins Tde1 and Tde2. Deletion and domain-swapping analysis identified that the C-terminal extension of VgrG1 specifically confers Tde1 secretion and Tde1-dependent interbacterial competition activity in planta, and the C-terminal variable region of VgrG2 governs this specificity for Tde2. Functional studies of VgrG1 and VgrG2 variants with stepwise deletion of the C terminus revealed that the C-terminal 31 aa (C31) of VgrG1 and 8 aa (C8) of VgrG2 are the molecular determinants specifically required for delivery of each cognate Tde toxin. Further in-depth studies on Tde toxin delivery mechanisms revealed that VgrG1 interacts with the adaptor/chaperone-effector complex (Tap-1-Tde1) in the absence of proline-alanine-alanine-arginine (PAAR) and the VgrG1-PAAR complex forms independent of Tap-1 and Tde1. Importantly, we identified the regions involved in these interactions. Although the entire C31 segment is required for binding with the Tap-1-Tde1 complex, only the first 15 aa of this region are necessary for PAAR binding. These results suggest that the VgrG1 C terminus interacts sequentially or simultaneously with the Tap-1-Tde1 complex and PAAR to govern Tde1 translocation across bacterial membranes and delivery into target cells for antibacterial activity.
Keywords: Agrobacterium tumefaciens; DNase effector; VgrG; interbacterial competition; type VI secretion system.
Conflict of interest statement
The authors declare no conflict of interest.
Figures








References
-
- Durand E, Cambillau C, Cascales E, Journet L. VgrG, Tae, Tle, and beyond: The versatile arsenal of Type VI secretion effectors. Trends Microbiol. 2014;22(9):498–507. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
