

Microbial Reconstitution Reverses Maternal Diet-Induced Social and Synaptic Deficits in Offspring
- PMID: 27315483
- PMCID: PMC5102250
- DOI: 10.1016/j.cell.2016.06.001
Microbial Reconstitution Reverses Maternal Diet-Induced Social and Synaptic Deficits in Offspring
Abstract
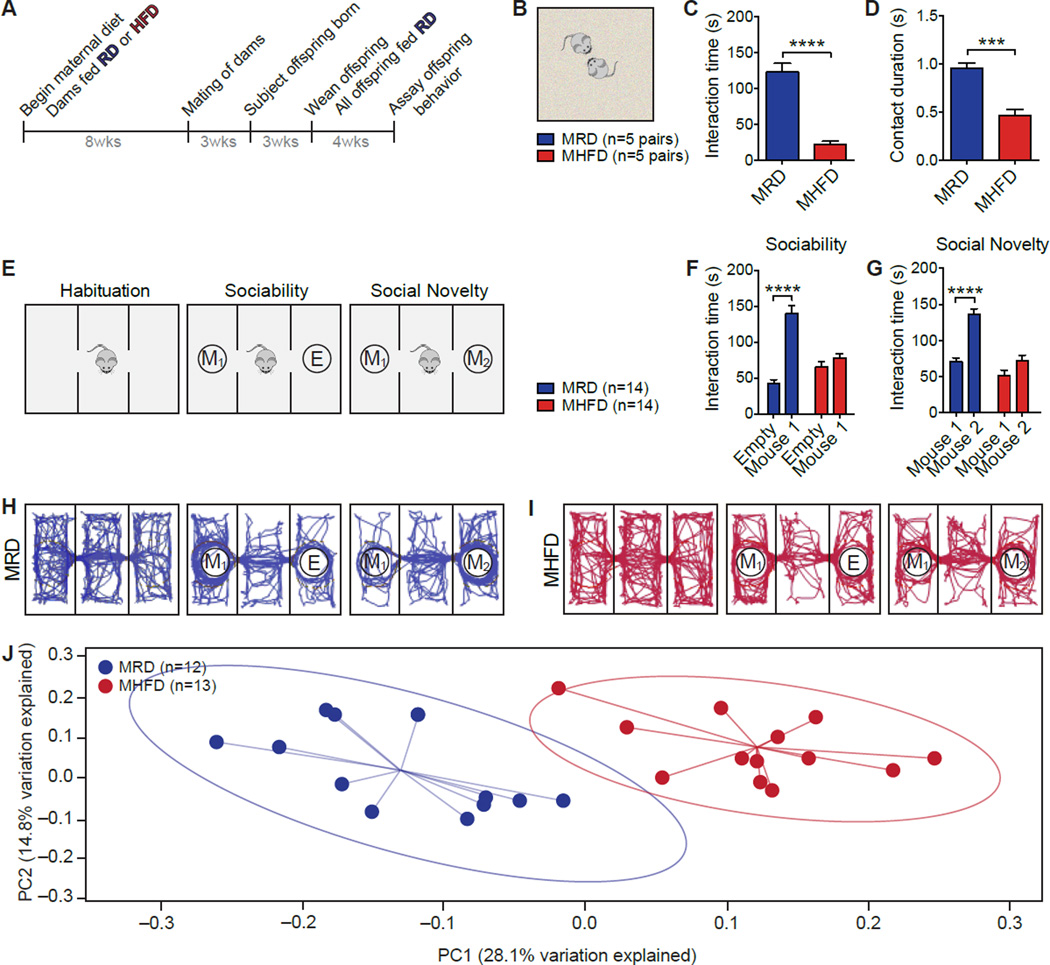
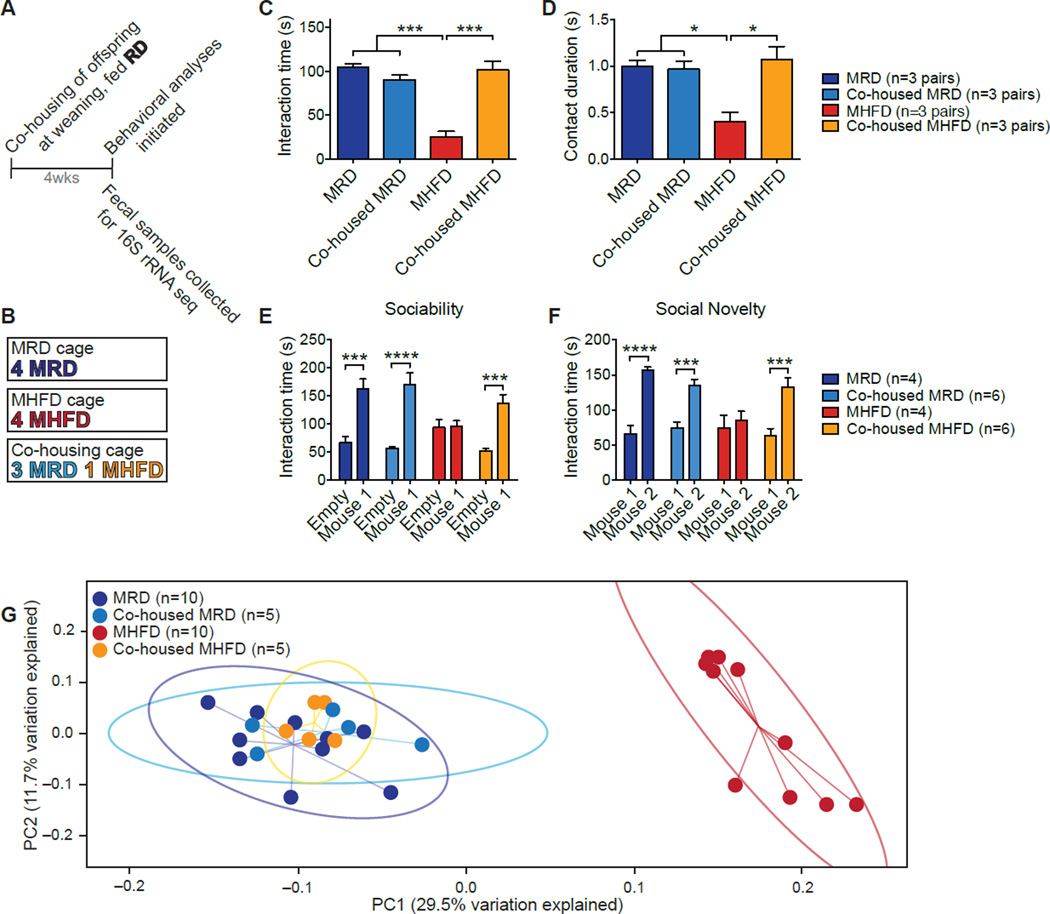
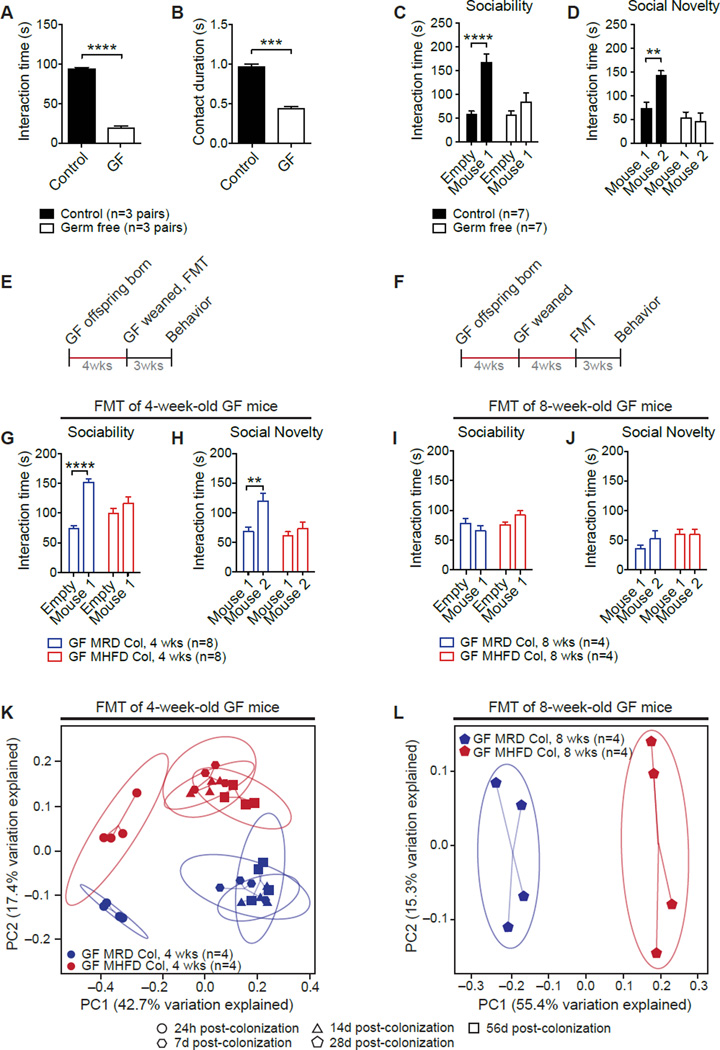
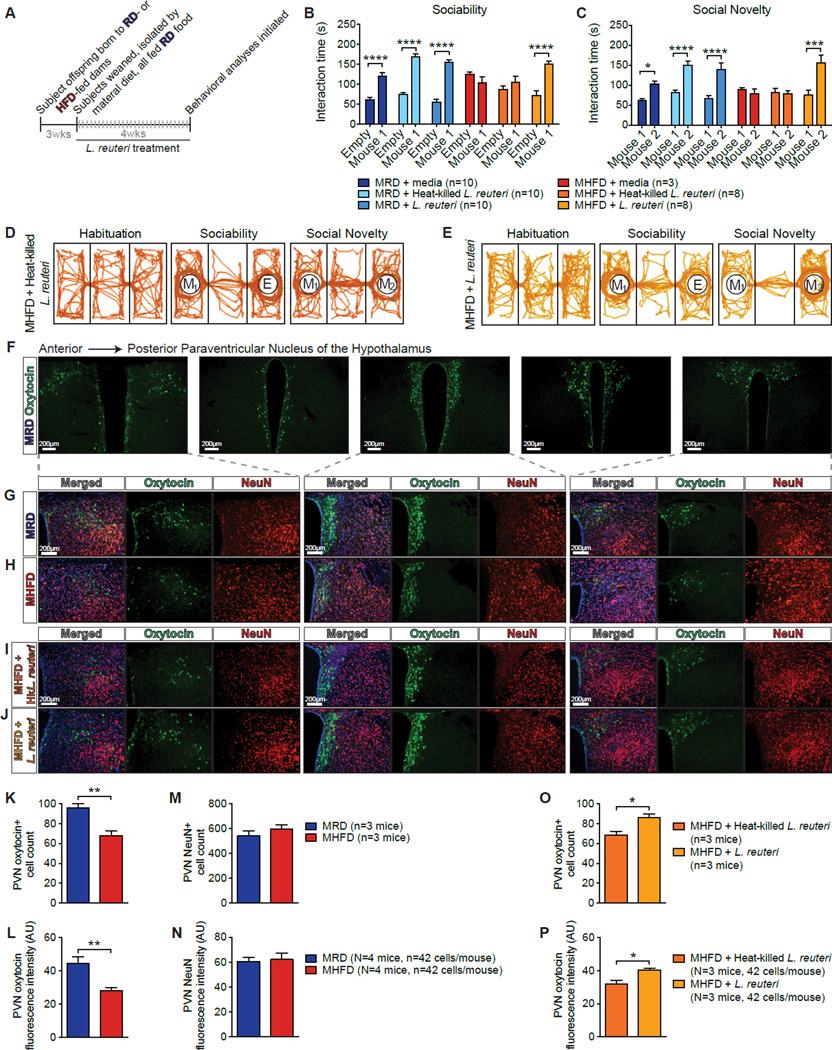
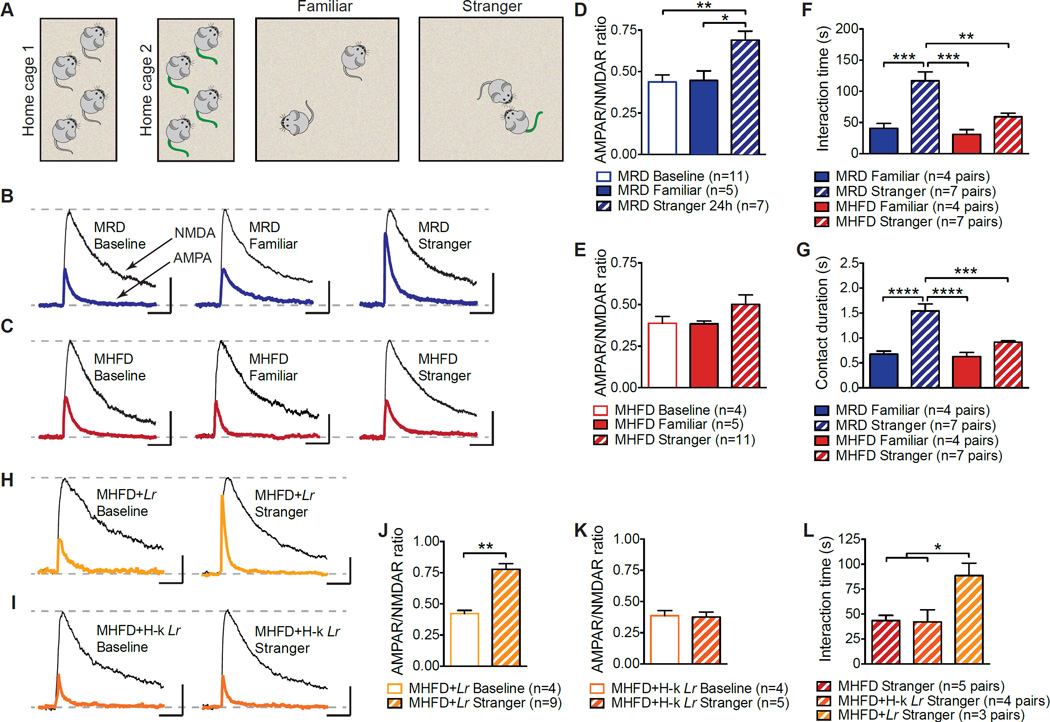
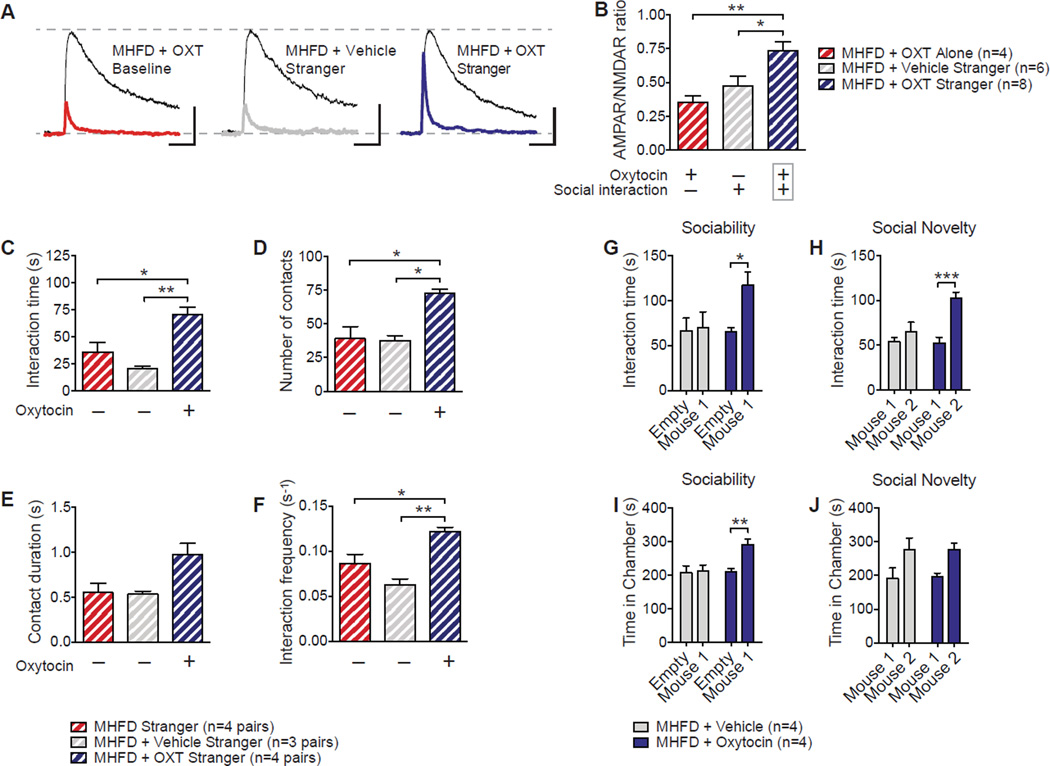
Maternal obesity during pregnancy has been associated with increased risk of neurodevelopmental disorders, including autism spectrum disorder (ASD), in offspring. Here, we report that maternal high-fat diet (MHFD) induces a shift in microbial ecology that negatively impacts offspring social behavior. Social deficits and gut microbiota dysbiosis in MHFD offspring are prevented by co-housing with offspring of mothers on a regular diet (MRD) and transferable to germ-free mice. In addition, social interaction induces synaptic potentiation (LTP) in the ventral tegmental area (VTA) of MRD, but not MHFD offspring. Moreover, MHFD offspring had fewer oxytocin immunoreactive neurons in the hypothalamus. Using metagenomics and precision microbiota reconstitution, we identified a single commensal strain that corrects oxytocin levels, LTP, and social deficits in MHFD offspring. Our findings causally link maternal diet, gut microbial imbalance, VTA plasticity, and behavior and suggest that probiotic treatment may relieve specific behavioral abnormalities associated with neurodevelopmental disorders. VIDEO ABSTRACT.
Keywords: autism; dysbiosis; high-fat diet (HFD); long-term potentiation (LTP); neurodevelopmental disorders; probiotic; ventral tegmental area (VTA).
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Gut-brain communication: Making friends with microbes.Nat Rev Neurosci. 2016 Sep;17(9):533. doi: 10.1038/nrn.2016.93. Epub 2016 Jul 7. Nat Rev Neurosci. 2016. PMID: 27383687 No abstract available.
-
Gut microbiota: Microbiota and behaviour: visiting the sins of the mother.Nat Rev Gastroenterol Hepatol. 2016 Sep;13(9):502-4. doi: 10.1038/nrgastro.2016.122. Epub 2016 Jul 27. Nat Rev Gastroenterol Hepatol. 2016. PMID: 27485788 No abstract available.
-
Defining 'good health'.Aging (Albany NY). 2016 Dec 29;8(12):3157-3158. doi: 10.18632/aging.101154. Aging (Albany NY). 2016. PMID: 28039492 Free PMC article. No abstract available.
References
-
- Benjamini Y, Drai D, Elmer G, Kafkafi N, Golani I. Controlling the false discovery rate in behavior genetics research. Behav Brain Res. 2001;125:279–284. - PubMed
-
- Bravo JA, Forsythe P, Chew MV, Escaravage E, Savignac HM, Dinan TG, Bienenstock J, Cryan JF. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:16050–16055. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
