

Cellular entry of the porcine epidemic diarrhea virus
- PMID: 27317167
- PMCID: PMC7114534
- DOI: 10.1016/j.virusres.2016.05.031
Cellular entry of the porcine epidemic diarrhea virus
Abstract
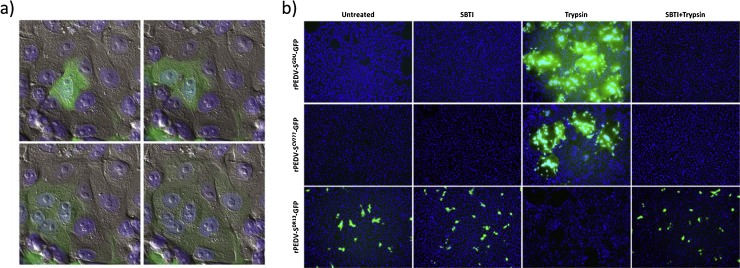
Porcine epidemic diarrhea virus (PEDV), a coronavirus discovered more than 40 years ago, regained notoriety recently by its devastating outbreaks in East Asia and the Americas, causing substantial economic losses to the swine husbandry. The virus replicates extensively and almost exclusively in the epithelial cells of the small intestine resulting in villus atrophy, malabsorption and severe diarrhea. Cellular entry of this enveloped virus is mediated by the large spike (S) glycoprotein, trimers of which mediate virus attachment to the target cell and subsequent membrane fusion. The S protein has a multidomain architecture and has been reported to bind to carbohydrate (sialic acid) and proteinaceous (aminopeptidase N) cell surface molecules. PEDV propagation in vitro requires the presence of trypsin(-like) proteases in the culture medium, which capacitates the fusion function of the S protein. Here we review the current data on PEDV entry into its host cell, including therein our new observations regarding the functional role of the sialic acid binding activity of the S protein in virus infection. Moreover, we summarize the recent progress on the proteolytic activation of PEDV S proteins, and discuss factors that may determine tissue tropism of PEDV in vivo.
Keywords: Coronavirus; Membrane fusion; PED; PEDV; Porcine epidemic diarrhea virus; Proteolytic activation; Receptor interaction; Sialic acid; Sialic acid binding; Spike; Virus; Virus entry; Virus tropism.
Copyright © 2016 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
Similar articles
-
Cell Attachment Domains of the Porcine Epidemic Diarrhea Virus Spike Protein Are Key Targets of Neutralizing Antibodies.J Virol. 2017 May 26;91(12):e00273-17. doi: 10.1128/JVI.00273-17. Print 2017 Jun 15. J Virol. 2017. PMID: 28381581 Free PMC article.
-
Three Main Inducers of Alphacoronavirus Infection of Enterocytes: Sialic Acid, Proteases, and Low pH.Intervirology. 2018;61(2):53-63. doi: 10.1159/000492424. Epub 2018 Sep 3. Intervirology. 2018. PMID: 30176660 Free PMC article. Review.
-
A Single Point Mutation Creating a Furin Cleavage Site in the Spike Protein Renders Porcine Epidemic Diarrhea Coronavirus Trypsin Independent for Cell Entry and Fusion.J Virol. 2015 Aug;89(15):8077-81. doi: 10.1128/JVI.00356-15. Epub 2015 May 13. J Virol. 2015. PMID: 25972540 Free PMC article.
-
Deciphering the biology of porcine epidemic diarrhea virus in the era of reverse genetics.Virus Res. 2016 Dec 2;226:152-171. doi: 10.1016/j.virusres.2016.05.003. Epub 2016 May 20. Virus Res. 2016. PMID: 27212685 Free PMC article. Review.
-
Porcine aminopeptidase N is a functional receptor for the PEDV coronavirus.Virology. 2007 Aug 15;365(1):166-72. doi: 10.1016/j.virol.2007.03.031. Epub 2007 Apr 30. Virology. 2007. PMID: 17467767 Free PMC article.
Cited by
-
The role of receptors in the cross-species spread of coronaviruses infecting humans and pigs.Arch Virol. 2024 Jan 24;169(2):35. doi: 10.1007/s00705-023-05956-7. Arch Virol. 2024. PMID: 38265497 Review.
-
Development of CRISPR-Mediated Nucleic Acid Detection Technologies and Their Applications in the Livestock Industry.Genes (Basel). 2022 Nov 2;13(11):2007. doi: 10.3390/genes13112007. Genes (Basel). 2022. PMID: 36360244 Free PMC article. Review.
-
Failure to experimentally infect 10 days-old piglets with a cell culture-propagated infectious stock of a classical genotype 1a porcine epidemic diarrhea virus.Front Vet Sci. 2023 Nov 16;10:1279162. doi: 10.3389/fvets.2023.1279162. eCollection 2023. Front Vet Sci. 2023. PMID: 38046573 Free PMC article.
-
Molecular Mechanism of Porcine Epidemic Diarrhea Virus Cell Tropism.mBio. 2022 Apr 26;13(2):e0373921. doi: 10.1128/mbio.03739-21. Epub 2022 Mar 14. mBio. 2022. PMID: 35285698 Free PMC article.
-
Clustered Regularly Interspaced Short Palindromic Repeat/Cas12a Mediated Multiplexable and Portable Detection Platform for GII Genotype Porcine Epidemic Diarrhoea Virus Rapid Diagnosis.Front Microbiol. 2022 Jun 9;13:920801. doi: 10.3389/fmicb.2022.920801. eCollection 2022. Front Microbiol. 2022. PMID: 35756009 Free PMC article.
References
-
- Baggen J., Thibaut H.J., Staring J., Jae L.T., Liu Y., Guo H., Slager J.J., de Bruin J.W., van Vliet A.L., Blomen V.A., Overduin P., Sheng J., de Haan Xander C.A., de Vries E., Meijer A., Rossmann M.G., Brummelkamp T.R., van Kuppeveld F.J. Enterovirus D68 receptor requirements unveiled by haploid genetics. Proc. Natl. Acad. Sci. U. S. A. 2016;113:1399–1404. - PMC - PubMed
-
- Chang S., Bae J., Kang T., Kim J., Chung G., Lim C., Laude H., Yang M., Jang Y. Identification of the epitope region capable of inducing neutralizing antibodies against the porcine epidemic diarrhea virus. Mol. Cells. 2002;14:295–299. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
