

mRNA capping: biological functions and applications
- PMID: 27317694
- PMCID: PMC5027499
- DOI: 10.1093/nar/gkw551
mRNA capping: biological functions and applications
Abstract
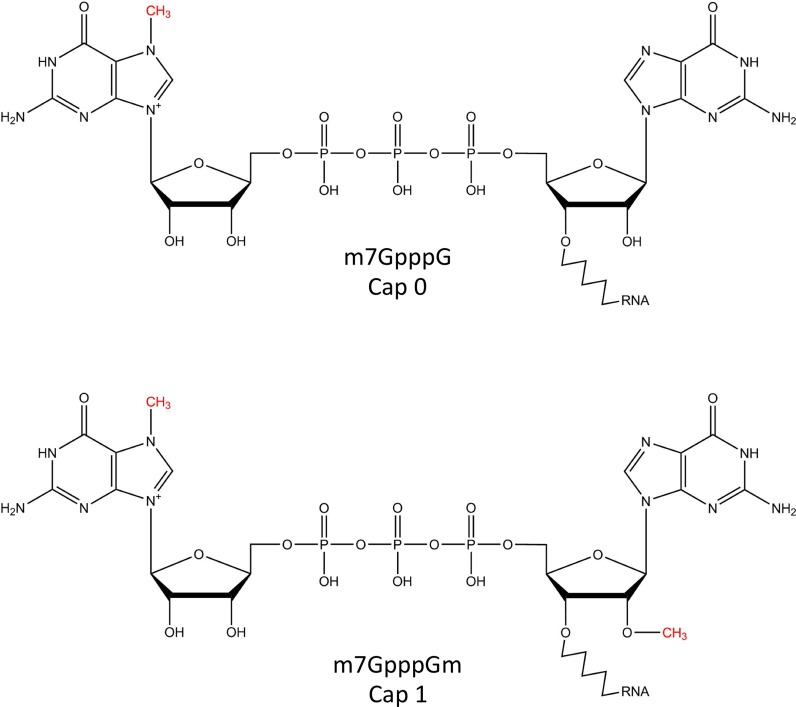
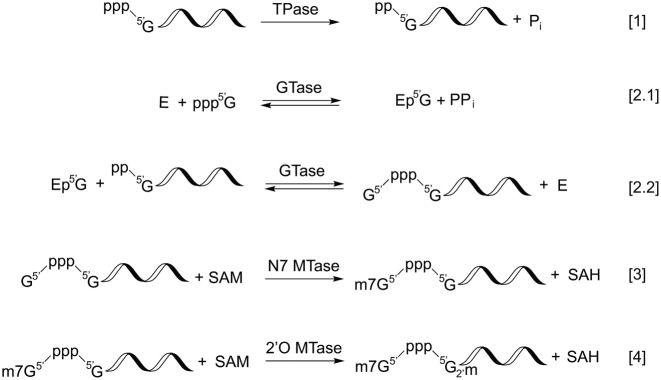
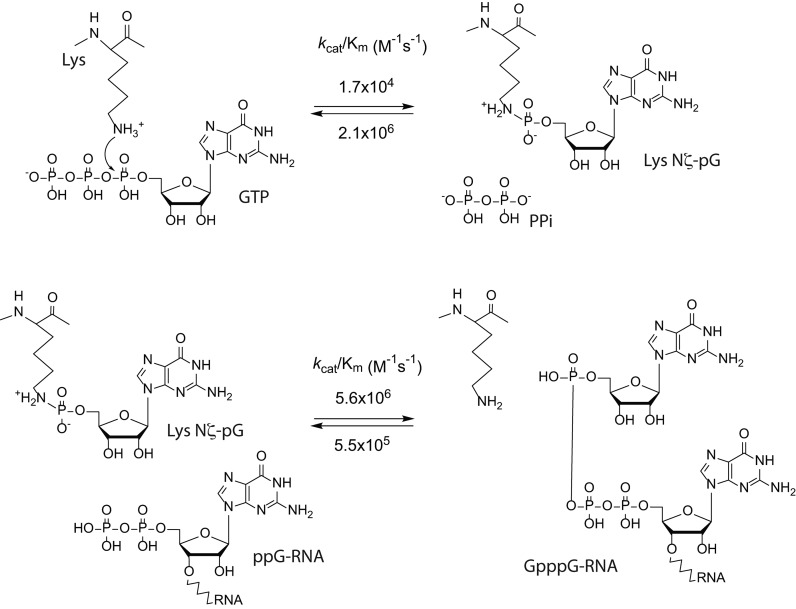
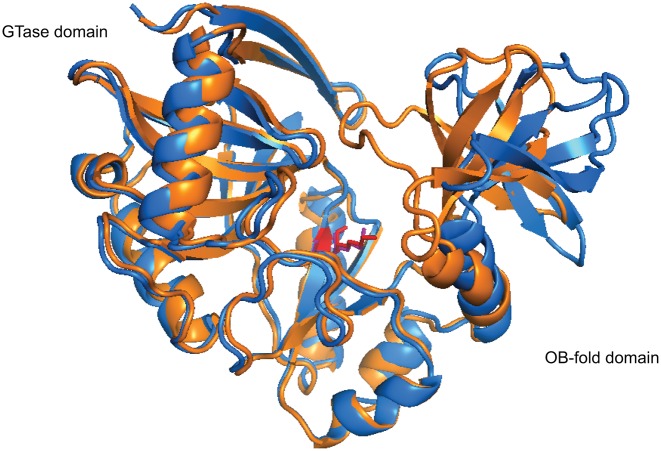
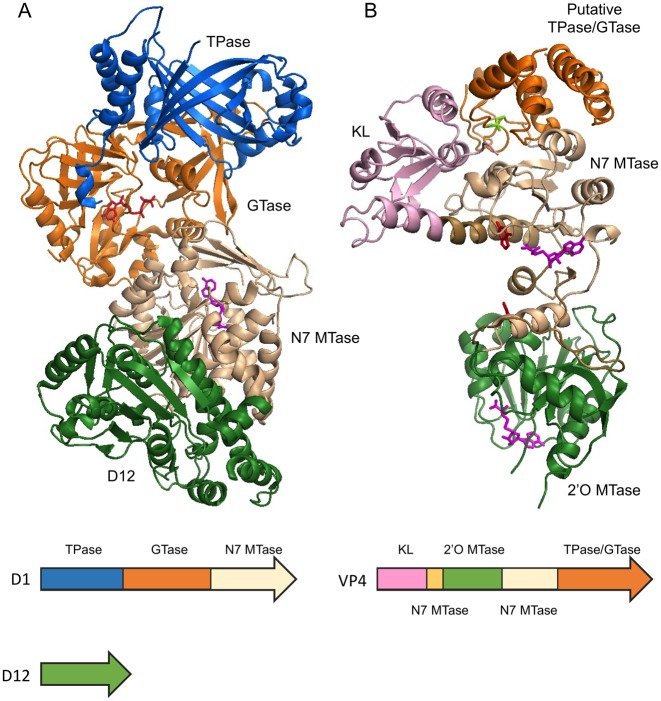
The 5' m7G cap is an evolutionarily conserved modification of eukaryotic mRNA. Decades of research have established that the m7G cap serves as a unique molecular module that recruits cellular proteins and mediates cap-related biological functions such as pre-mRNA processing, nuclear export and cap-dependent protein synthesis. Only recently has the role of the cap 2'O methylation as an identifier of self RNA in the innate immune system against foreign RNA has become clear. The discovery of the cytoplasmic capping machinery suggests a novel level of control network. These new findings underscore the importance of a proper cap structure in the synthesis of functional messenger RNA. In this review, we will summarize the current knowledge of the biological roles of mRNA caps in eukaryotic cells. We will also discuss different means that viruses and their host cells use to cap their RNA and the application of these capping machineries to synthesize functional mRNA. Novel applications of RNA capping enzymes in the discovery of new RNA species and sequencing the microbiome transcriptome will also be discussed. We will end with a summary of novel findings in RNA capping and the questions these findings pose.
© The Author(s) 2016. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
