

Functional analysis of AEBP2, a PRC2 Polycomb protein, reveals a Trithorax phenotype in embryonic development and in ESCs
- PMID: 27317809
- PMCID: PMC5004903
- DOI: 10.1242/dev.123935
Functional analysis of AEBP2, a PRC2 Polycomb protein, reveals a Trithorax phenotype in embryonic development and in ESCs
Abstract
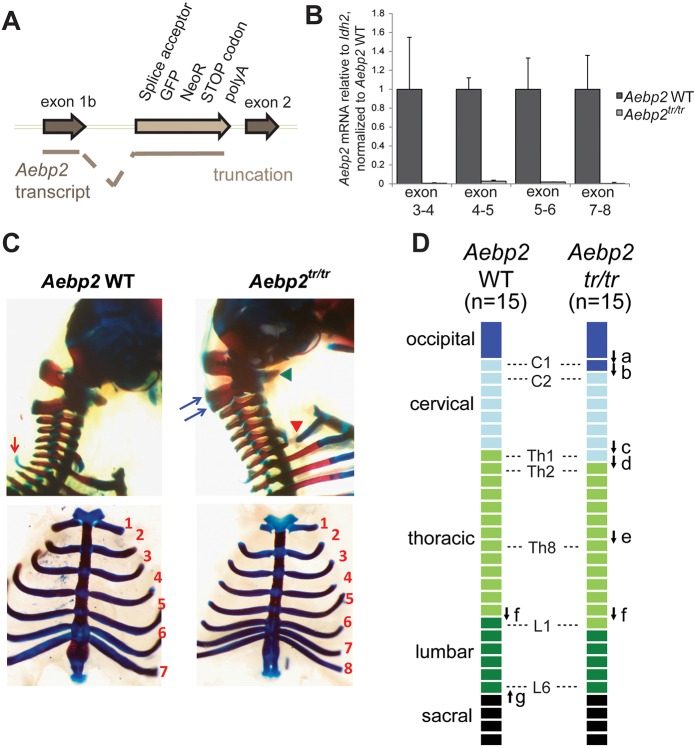
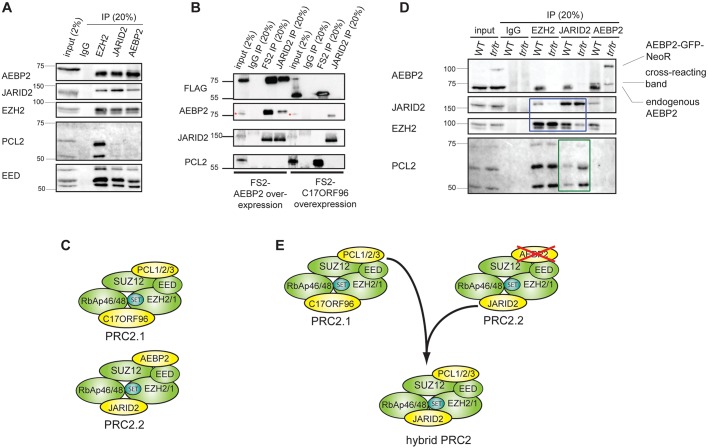
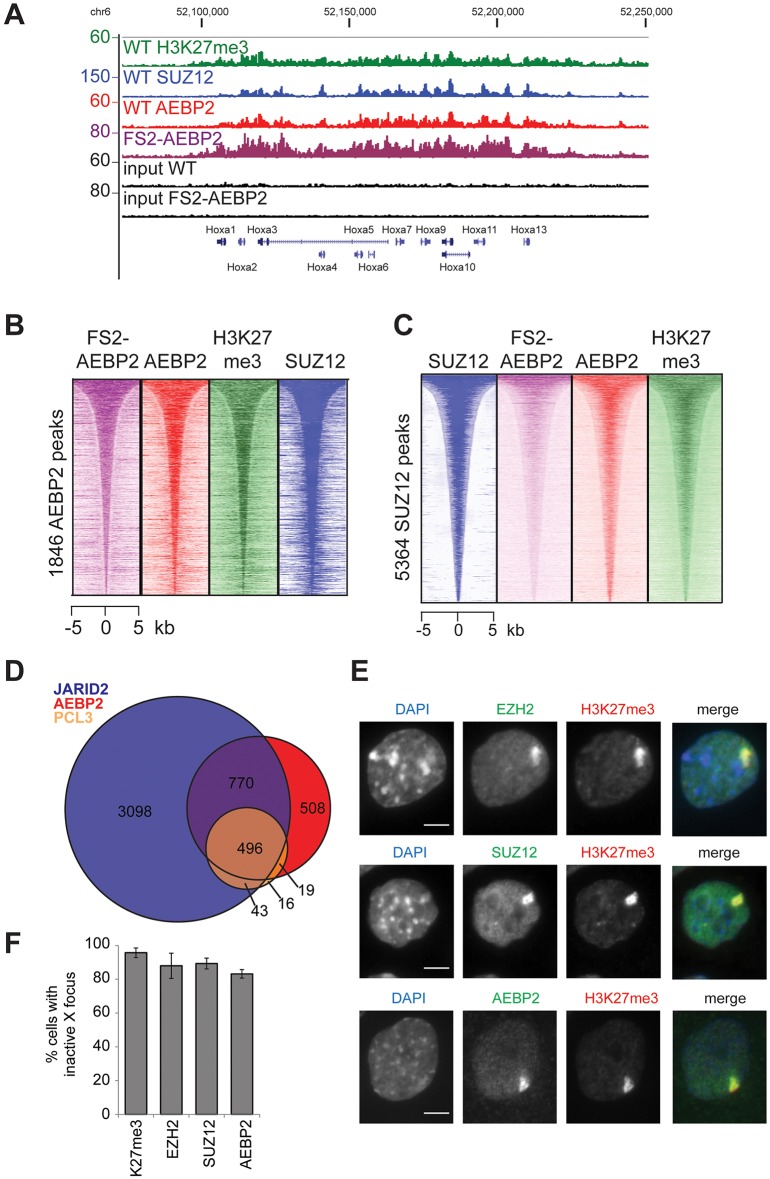
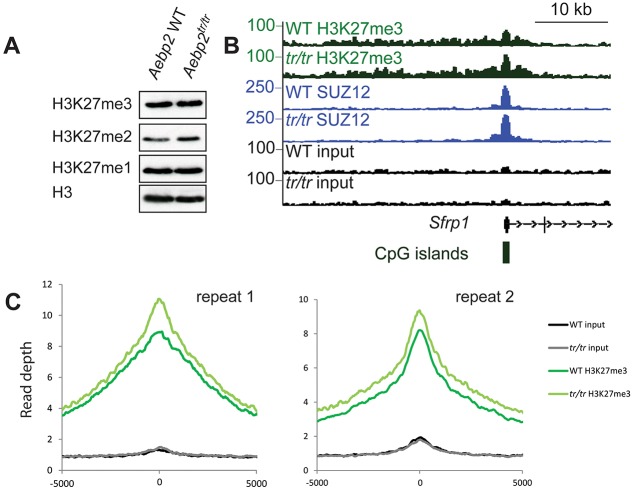
The Polycomb repressive complexes PRC1 and PRC2 are key mediators of heritable gene silencing in multicellular organisms. Here, we characterise AEBP2, a known PRC2 co-factor which, in vitro, has been shown to stimulate PRC2 activity. We show that AEBP2 localises specifically to PRC2 target loci, including the inactive X chromosome. Proteomic analysis confirms that AEBP2 associates exclusively with PRC2 complexes. However, analysis of embryos homozygous for a targeted mutation of Aebp2 unexpectedly revealed a Trithorax phenotype, normally linked to antagonism of Polycomb function. Consistent with this, we observe elevated levels of PRC2-mediated histone H3K27 methylation at target loci in Aebp2 mutant embryonic stem cells (ESCs). We further demonstrate that mutant ESCs assemble atypical hybrid PRC2 subcomplexes, potentially accounting for enhancement of Polycomb activity, and suggesting that AEBP2 normally plays a role in defining the mutually exclusive composition of PRC2 subcomplexes.
Keywords: AEBP2; Chromatin; Mouse; PRC2; Polycomb; Trithorax.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
-
- Akasaka T., Kanno M., Balling R., Mieza M. A., Taniguchi M. and Koseki H. (1996). A role for mel-18, a Polycomb group-related vertebrate gene, during the anteroposterior specification of the axial skeleton. Development 122, 1513-1522. - PubMed
-
- Blackledge N. P., Farcas A. M., Kondo T., King H. W., McGouran J. F., Hanssen L. L. P., Ito S., Cooper S., Kondo K., Koseki Y. et al. (2014). Variant PRC1 complex-dependent H2A ubiquitylation drives PRC2 recruitment and polycomb domain formation. Cell 157, 1445-1459. 10.1016/j.cell.2014.05.004 - DOI - PMC - PubMed
-
- Brien G. L., Gambero G., O'Connell D. J., Jerman E., Turner S. A., Egan C. M., Dunne E. J., Jurgens M. C., Wynne K., Piao L. et al. (2012). Polycomb PHF19 binds H3K36me3 and recruits PRC2 and demethylase NO66 to embryonic stem cell genes during differentiation. Nat. Struct. Mol. Biol. 19, 1273-1281. 10.1038/nsmb.2449 - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
