

Loss of the RNA-binding protein TACO1 causes late-onset mitochondrial dysfunction in mice
- PMID: 27319982
- PMCID: PMC4915168
- DOI: 10.1038/ncomms11884
Loss of the RNA-binding protein TACO1 causes late-onset mitochondrial dysfunction in mice
Abstract
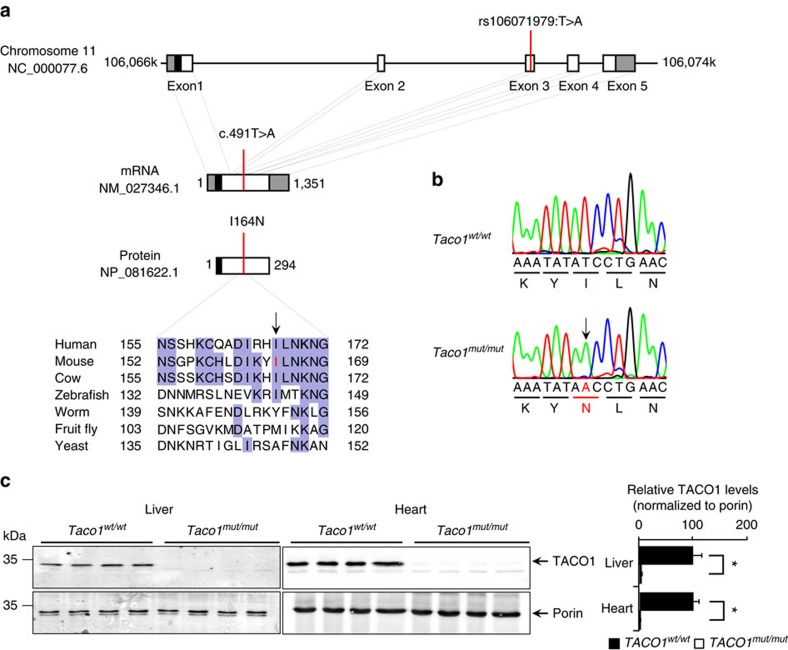
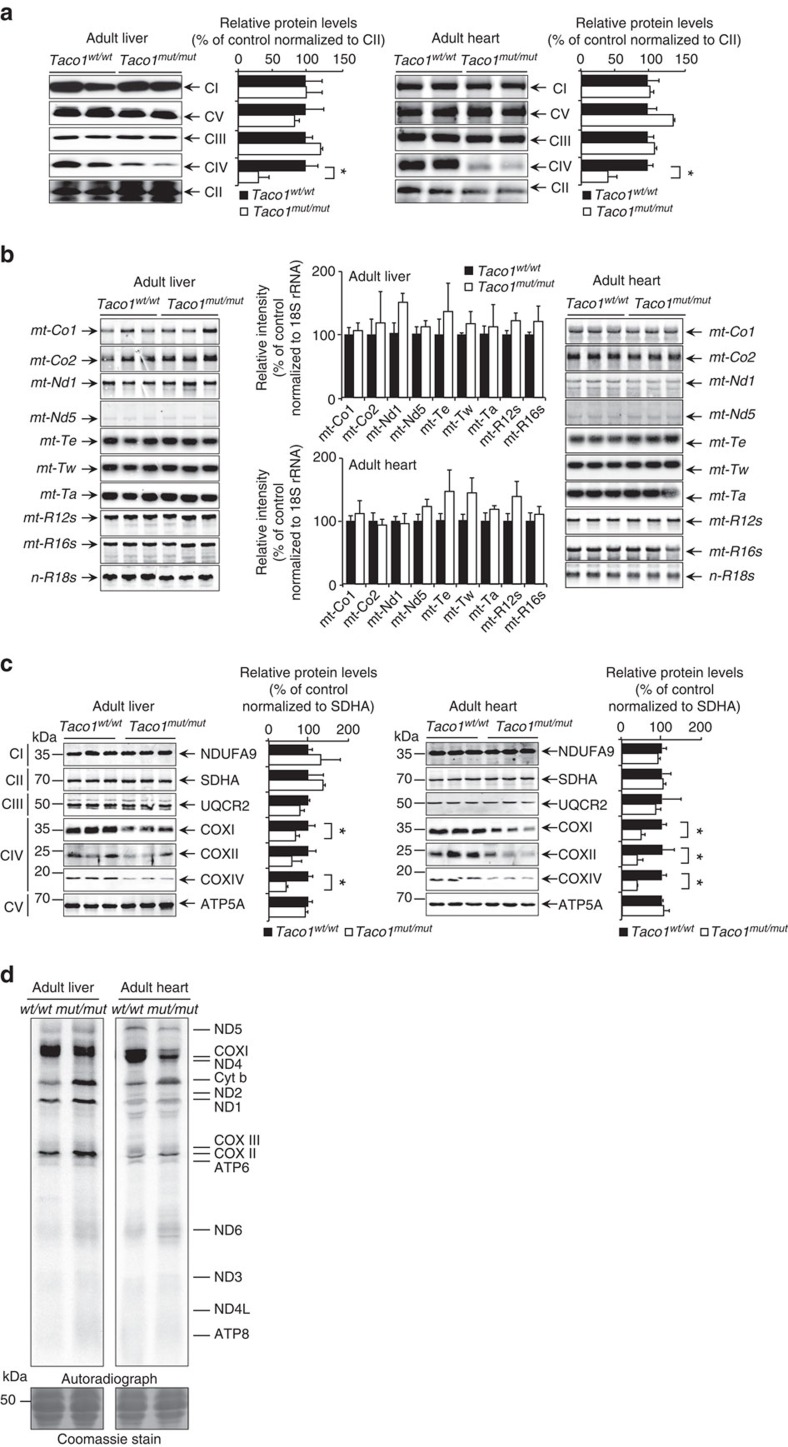
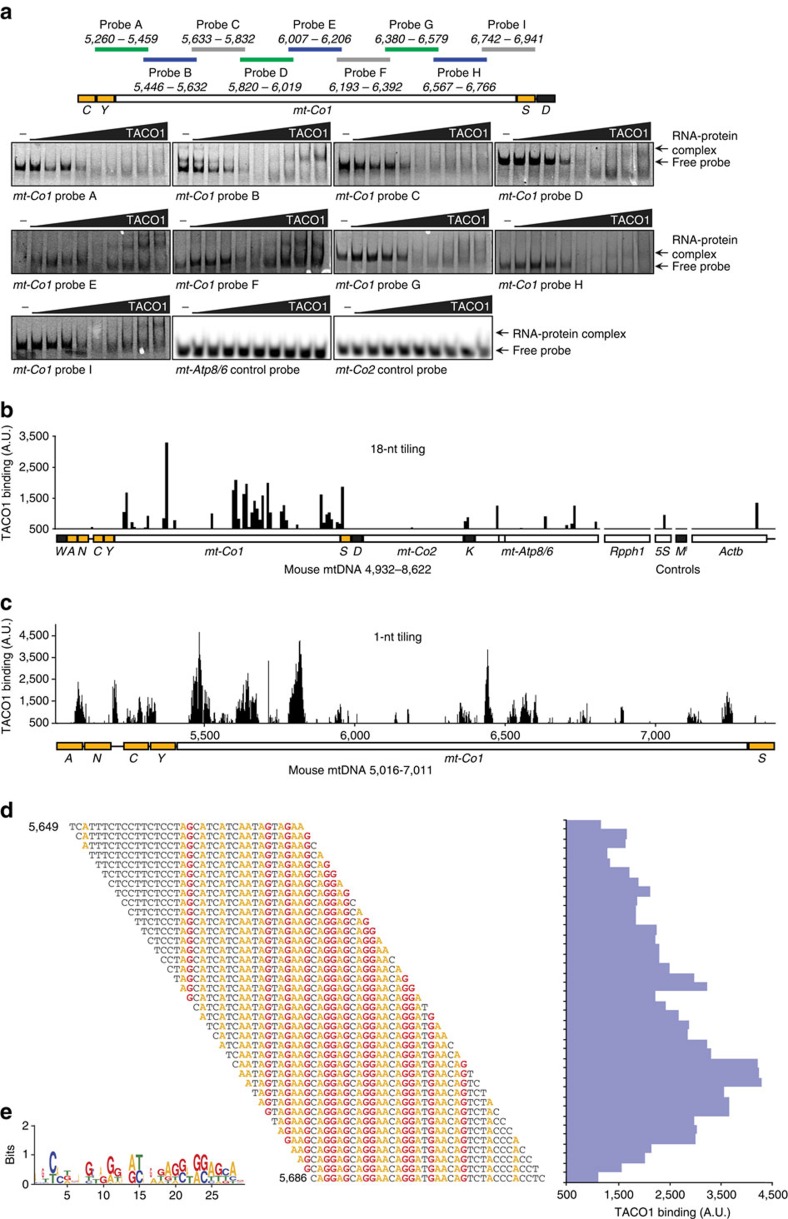
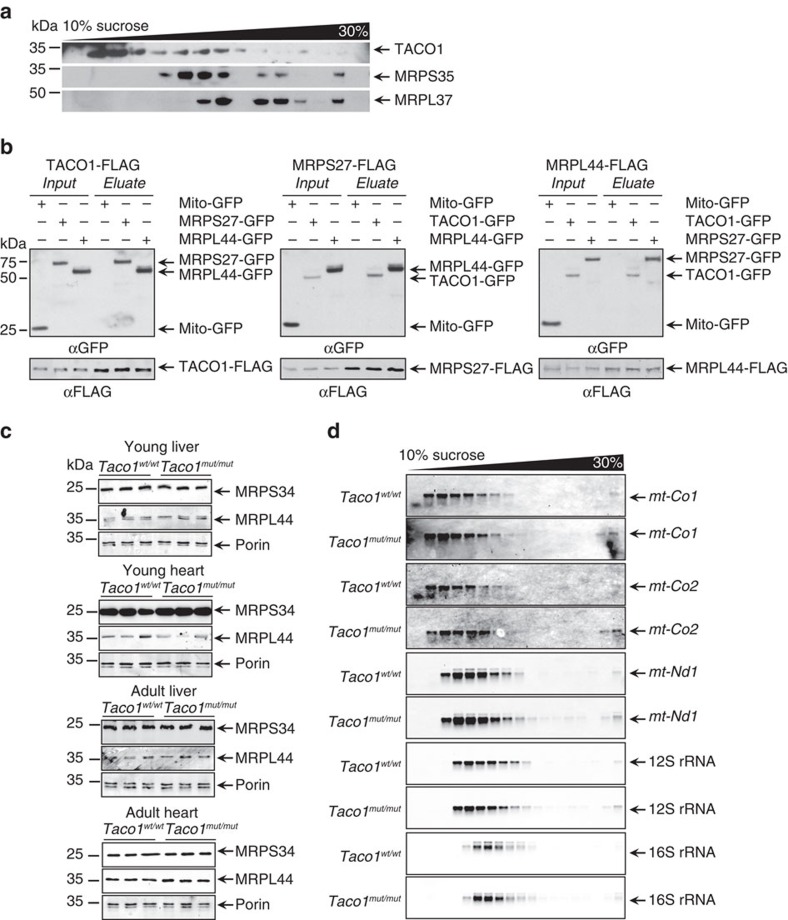
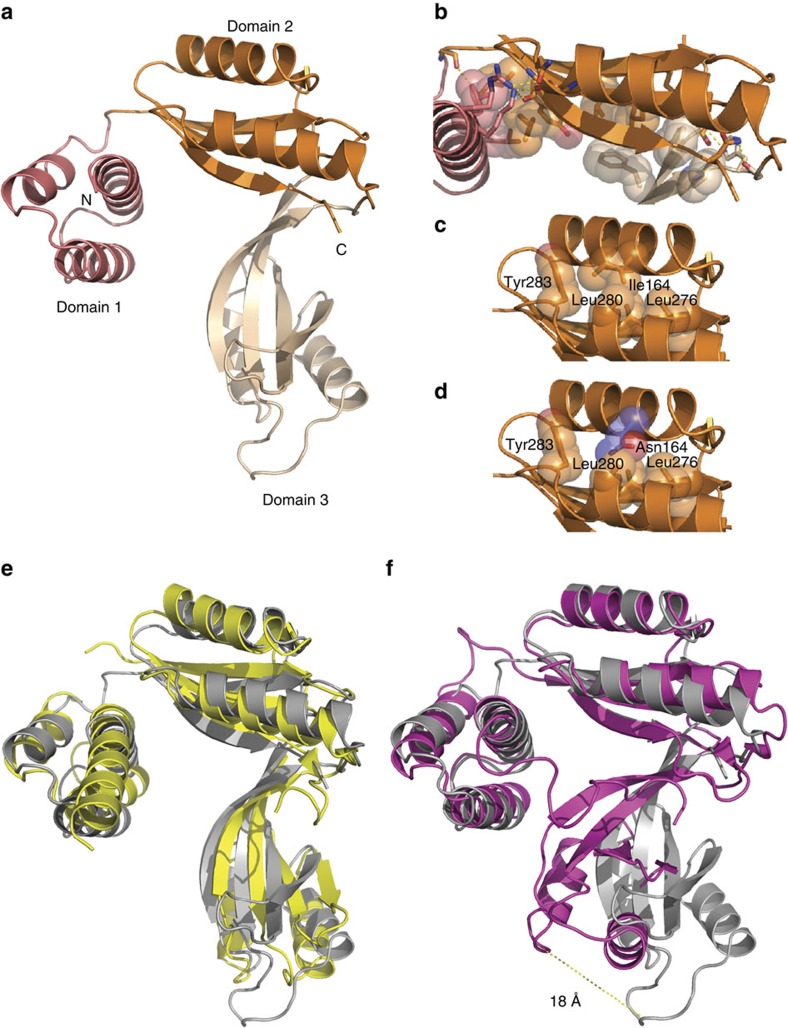
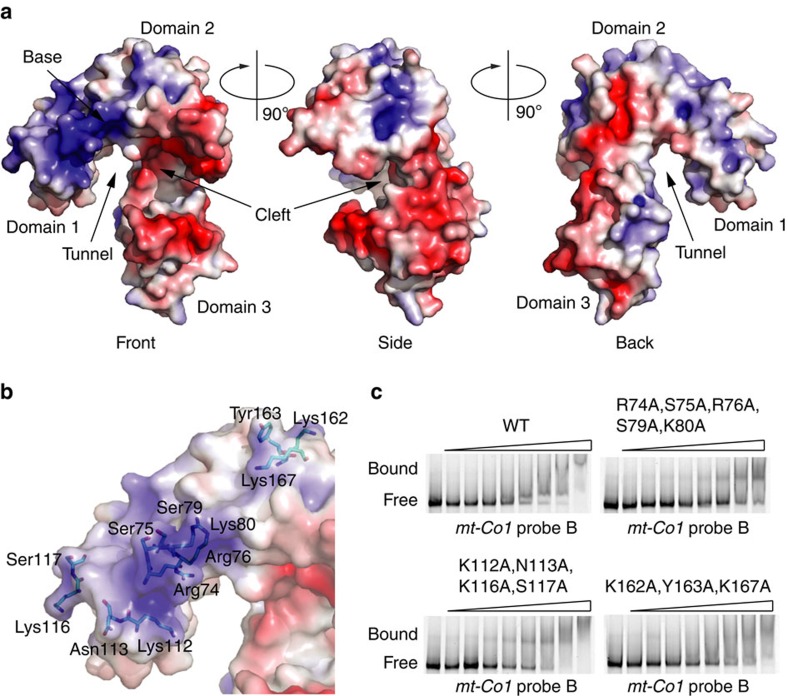
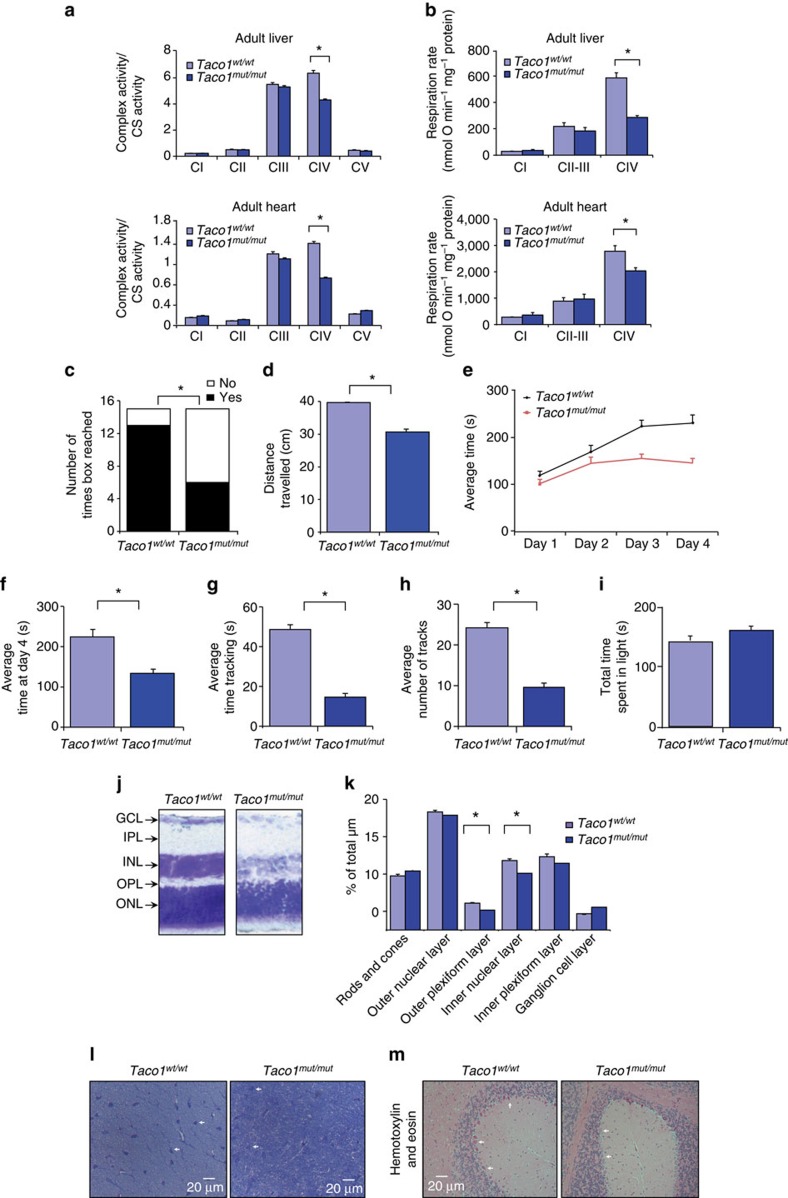
The recognition and translation of mammalian mitochondrial mRNAs are poorly understood. To gain further insights into these processes in vivo, we characterized mice with a missense mutation that causes loss of the translational activator of cytochrome oxidase subunit I (TACO1). We report that TACO1 is not required for embryonic survival, although the mutant mice have substantially reduced COXI protein, causing an isolated complex IV deficiency. We show that TACO1 specifically binds the mt-Co1 mRNA and is required for translation of COXI through its association with the mitochondrial ribosome. We determined the atomic structure of TACO1, revealing three domains in the shape of a hook with a tunnel between domains 1 and 3. Mutations in the positively charged domain 1 reduce RNA binding by TACO1. The Taco1 mutant mice develop a late-onset visual impairment, motor dysfunction and cardiac hypertrophy and thus provide a useful model for future treatment trials for mitochondrial disease.
Figures
References
-
- Hallberg B. M. & Larsson N.-G. Making proteins in the powerhouse. Cell Metab. 20, 226–240 (2014). - PubMed
-
- Fox T. Translational control of endogenous and recoded nuclear genes in yeast mitochondria: regulation and membrane targeting. Experientia 52, 1130–1135 (1996). - PubMed
-
- Small I. D., Rackham O. & Filipovska A. Organelle transcriptomes: products of a deconstructed genome. Curr. Opin. Microbiol. 16, 652–658 (2013). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
