

Hepatic cytochromes P450: structural degrons and barcodes, posttranslational modifications and cellular adapters in the ERAD-endgame
- PMID: 27320797
- PMCID: PMC5094896
- DOI: 10.1080/03602532.2016.1195403
Hepatic cytochromes P450: structural degrons and barcodes, posttranslational modifications and cellular adapters in the ERAD-endgame
Abstract
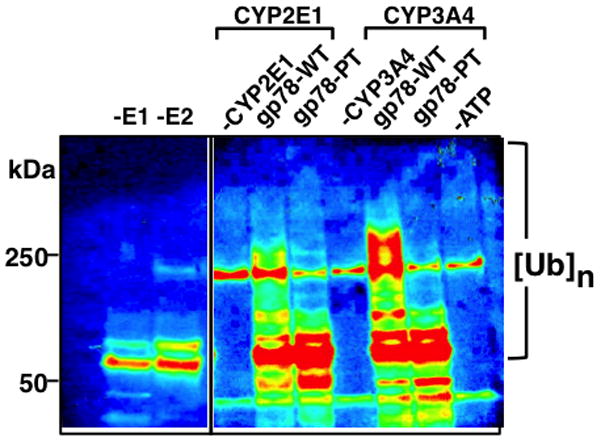
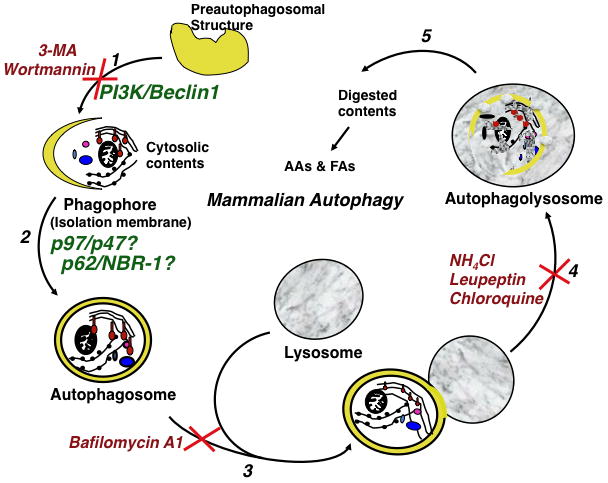
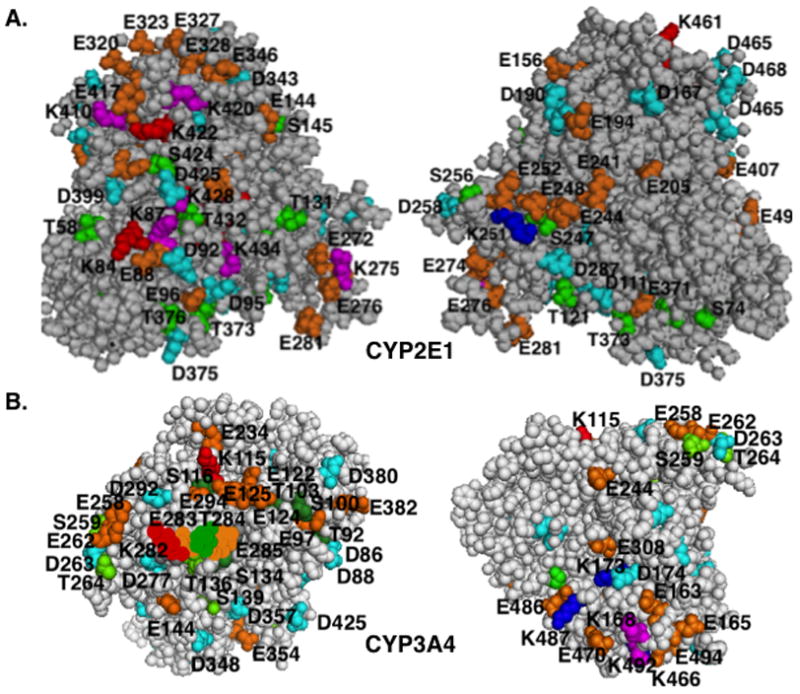
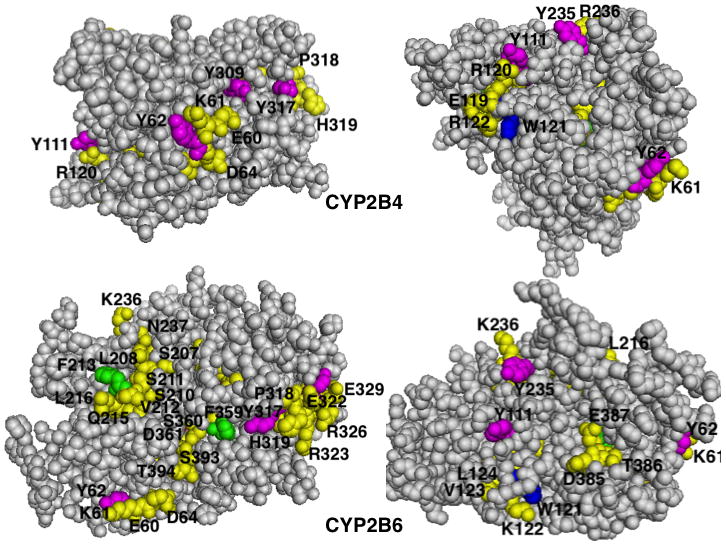
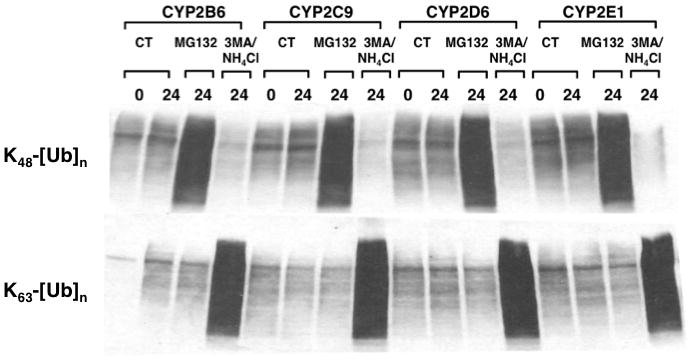
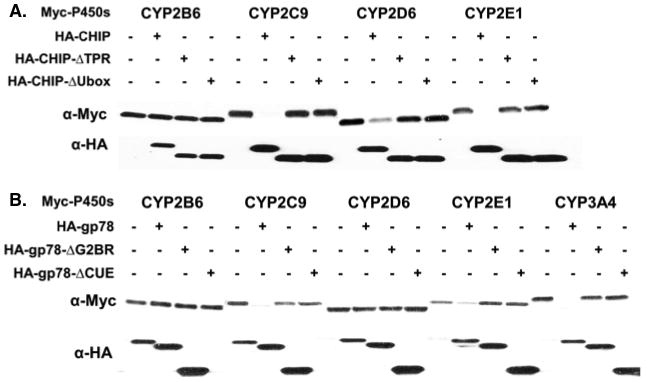
The endoplasmic reticulum (ER)-anchored hepatic cytochromes P450 (P450s) are enzymes that metabolize endo- and xenobiotics i.e. drugs, carcinogens, toxins, natural and chemical products. These agents modulate liver P450 content through increased synthesis or reduction via inactivation and/or proteolytic degradation, resulting in clinically significant drug-drug interactions. P450 proteolytic degradation occurs via ER-associated degradation (ERAD) involving either of two distinct routes: Ubiquitin (Ub)-dependent 26S proteasomal degradation (ERAD/UPD) or autophagic lysosomal degradation (ERAD/ALD). CYP3A4, the major human liver/intestinal P450, and the fast-turnover CYP2E1 species are degraded via ERAD/UPD entailing multisite protein phosphorylation and subsequent ubiquitination by gp78 and CHIP E3 Ub-ligases. We are gaining insight into the nature of the structural determinants involved in CYP3A4 and CYP2E1 molecular recognition in ERAD/UPD [i.e. K48-linked polyUb chains and linear and/or "conformational" phosphodegrons consisting either of consecutive sequences on surface loops and/or disordered regions, or structurally-assembled surface clusters of negatively charged acidic (Asp/Glu) and phosphorylated (Ser/Thr) residues, within or vicinal to which, Lys-residues are targeted for ubiquitination]. Structural inspection of select human liver P450s reveals that such linear or conformational phosphodegrons may indeed be a common P450-ERAD/UPD feature. By contrast, although many P450s such as the slow-turnover CYP2E1 species and rat liver CYP2B1 and CYP2C11 are degraded via ERAD/ALD, little is known about the mechanism of their ALD-targeting. On the basis of our current knowledge of ALD-substrate targeting, we propose a tripartite conjunction of K63-linked Ub-chains, P450 structural "LIR" motifs and selective cellular "cargo receptors" as plausible P450-ALD determinants.
Keywords: 26S proteasome; CHIP; Cytochromes P450; E2/E3 ubiquitin ligases; ERAD; K48 & K63 ubiquitination; autophagic lysosomal degradation; gp78 E3; p62; p97; phosphodegrons.
Conflict of interest statement
No interests to declare.
Figures







Similar articles
-
Induction via Functional Protein Stabilization of Hepatic Cytochromes P450 upon gp78/Autocrine Motility Factor Receptor (AMFR) Ubiquitin E3-Ligase Genetic Ablation in Mice: Therapeutic and Toxicological Relevance.Mol Pharmacol. 2019 Nov;96(5):641-654. doi: 10.1124/mol.119.117069. Epub 2019 Sep 6. Mol Pharmacol. 2019. PMID: 31492698 Free PMC article.
-
Human liver cytochrome P450 3A4 ubiquitination: molecular recognition by UBC7-gp78 autocrine motility factor receptor and UbcH5a-CHIP-Hsc70-Hsp40 E2-E3 ubiquitin ligase complexes.J Biol Chem. 2015 Feb 6;290(6):3308-32. doi: 10.1074/jbc.M114.611525. Epub 2014 Dec 1. J Biol Chem. 2015. PMID: 25451919 Free PMC article.
-
Hepatic cytochrome P450 ubiquitination: conformational phosphodegrons for E2/E3 recognition?IUBMB Life. 2014 Feb;66(2):78-88. doi: 10.1002/iub.1247. Epub 2014 Feb 3. IUBMB Life. 2014. PMID: 24488826 Free PMC article. Review.
-
Multisite phosphorylation of human liver cytochrome P450 3A4 enhances Its gp78- and CHIP-mediated ubiquitination: a pivotal role of its Ser-478 residue in the gp78-catalyzed reaction.Mol Cell Proteomics. 2012 Feb;11(2):M111.010132. doi: 10.1074/mcp.M111.010132. Epub 2011 Nov 17. Mol Cell Proteomics. 2012. PMID: 22101235 Free PMC article.
-
Cytochrome P450 endoplasmic reticulum-associated degradation (ERAD): therapeutic and pathophysiological implications.Acta Pharm Sin B. 2020 Jan;10(1):42-60. doi: 10.1016/j.apsb.2019.11.002. Epub 2019 Nov 8. Acta Pharm Sin B. 2020. PMID: 31993306 Free PMC article. Review.
Cited by
-
Progesterone receptor membrane component 1 (PGRMC1) binds and stabilizes cytochromes P450 through a heme-independent mechanism.J Biol Chem. 2021 Nov;297(5):101316. doi: 10.1016/j.jbc.2021.101316. Epub 2021 Oct 20. J Biol Chem. 2021. PMID: 34678314 Free PMC article.
-
Induction by Phenobarbital of Phase I and II Xenobiotic-Metabolizing Enzymes in Bovine Liver: An Overall Catalytic and Immunochemical Characterization.Int J Mol Sci. 2022 Mar 24;23(7):3564. doi: 10.3390/ijms23073564. Int J Mol Sci. 2022. PMID: 35408925 Free PMC article.
-
Time-dependent enzyme inactivation: Numerical analyses of in vitro data and prediction of drug-drug interactions.Pharmacol Ther. 2020 Feb;206:107449. doi: 10.1016/j.pharmthera.2019.107449. Epub 2019 Dec 11. Pharmacol Ther. 2020. PMID: 31836452 Free PMC article. Review.
-
Post-transcriptional Regulation of UGT2B10 Hepatic Expression and Activity by Alternative Splicing.Drug Metab Dispos. 2018 May;46(5):514-524. doi: 10.1124/dmd.117.079921. Epub 2018 Feb 9. Drug Metab Dispos. 2018. PMID: 29438977 Free PMC article.
-
Protein degradation: expanding the toolbox to restrain cancer drug resistance.J Hematol Oncol. 2023 Jan 24;16(1):6. doi: 10.1186/s13045-023-01398-5. J Hematol Oncol. 2023. PMID: 36694209 Free PMC article. Review.
References
-
- Aguiar M, Masse R, Gibbs BF. Regulation of cytochrome P450 by posttranslational modification. Drug Metab Rev. 2005;37:379–404. - PubMed
-
- Ahner A, Brodsky JL. Checkpoints in ER-associated degradation: excuse me, which way to the proteasome? Trends Cell Biol. 2004;14:474–478. - PubMed
-
- Anderson DJ, Le Moigne R, Djakovic S, Kumar B, Rice J, Wong S, Wang J, Yao B, Valle E, Kiss von Soly S, Madriaga A, Soriano F, Menon MK, Wu ZY, Kampmann M, Chen Y, Weissman JS, Aftab BT, Yakes FM, Shawver L, Zhou HJ, Wustrow D, Rolfe M. Targeting the AAA ATPase p97 as an Approach to Treat Cancer through Disruption of Protein Homeostasis. Cancer Cell. 2015;28:653–665. - PMC - PubMed
-
- Bandiera S, Weidlich S, Harth V, Broede P, Ko Y, Friedberg T. Proteasomal degradation of human CYP1B1: effect of the Asn453Ser polymorphism on the post-translational regulation of CYP1B1 expression. Mol Pharmacol. 2005;67:435–443. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources