

Hepatic Fatty Acid Oxidation Restrains Systemic Catabolism during Starvation
- PMID: 27320917
- PMCID: PMC4927362
- DOI: 10.1016/j.celrep.2016.05.062
Hepatic Fatty Acid Oxidation Restrains Systemic Catabolism during Starvation
Abstract
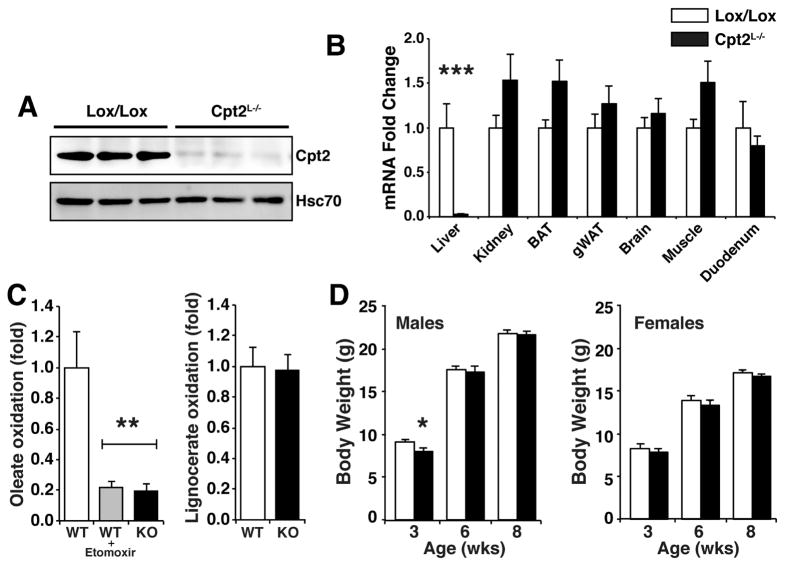
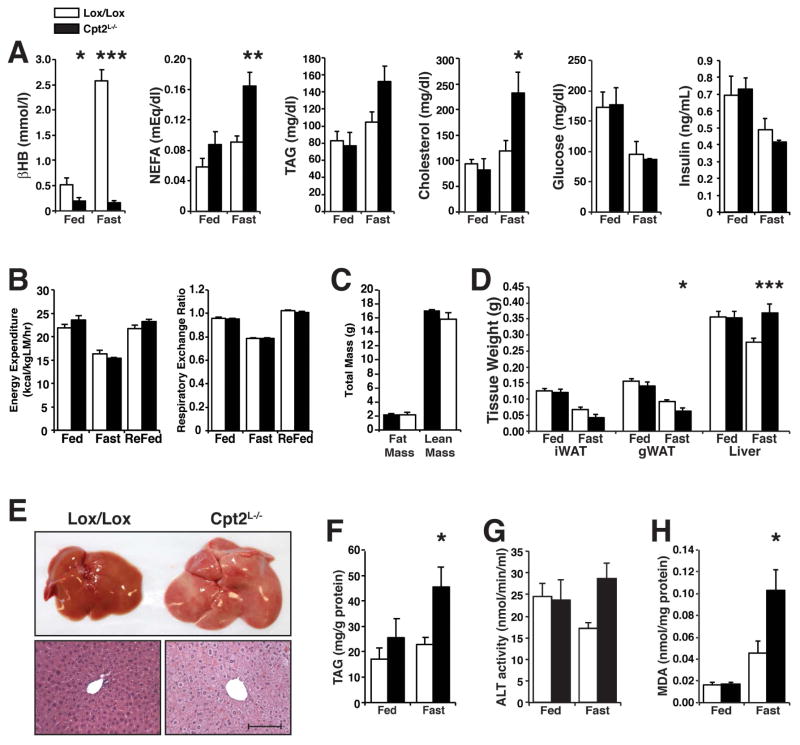
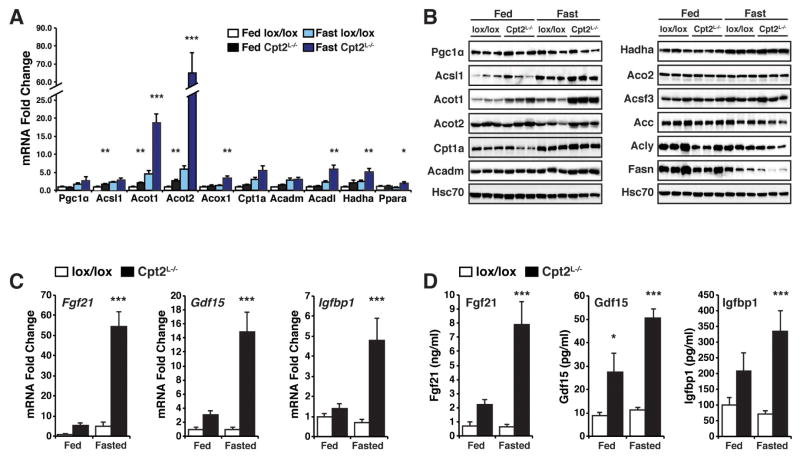
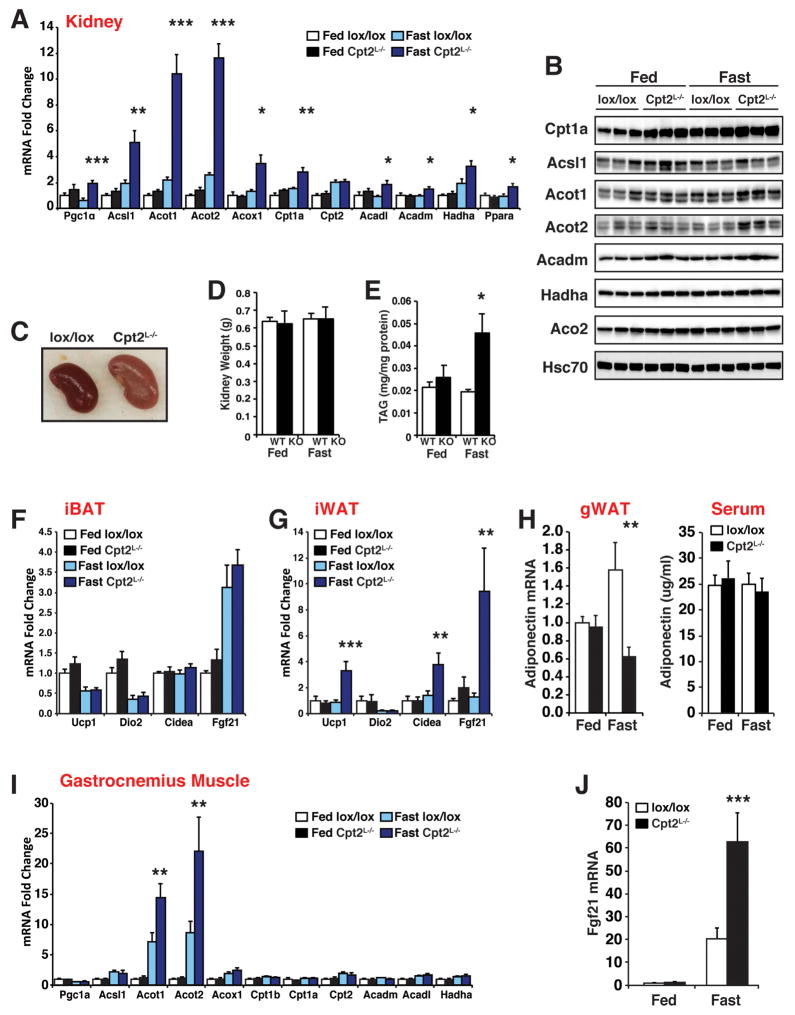
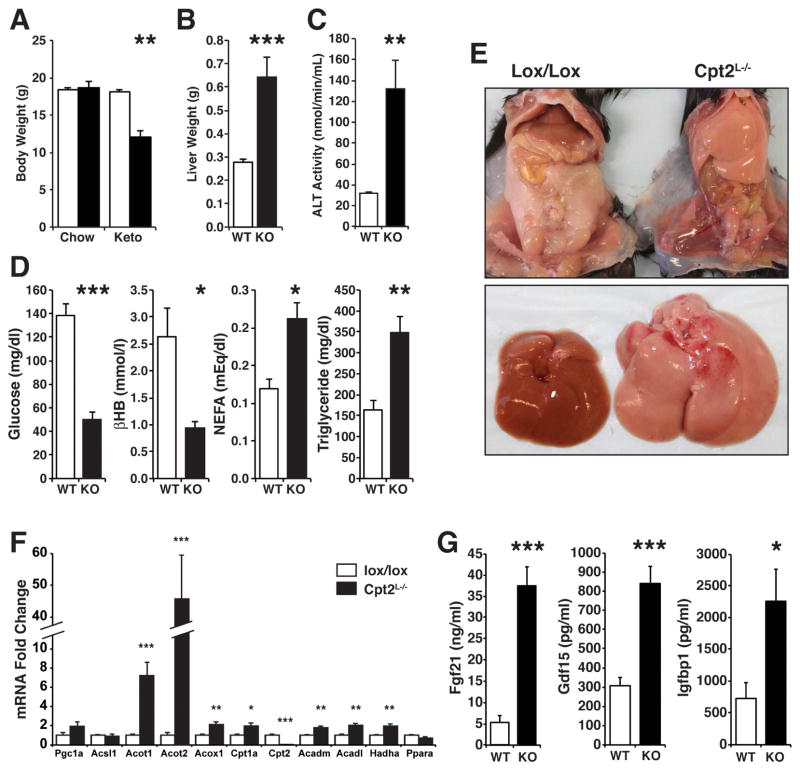
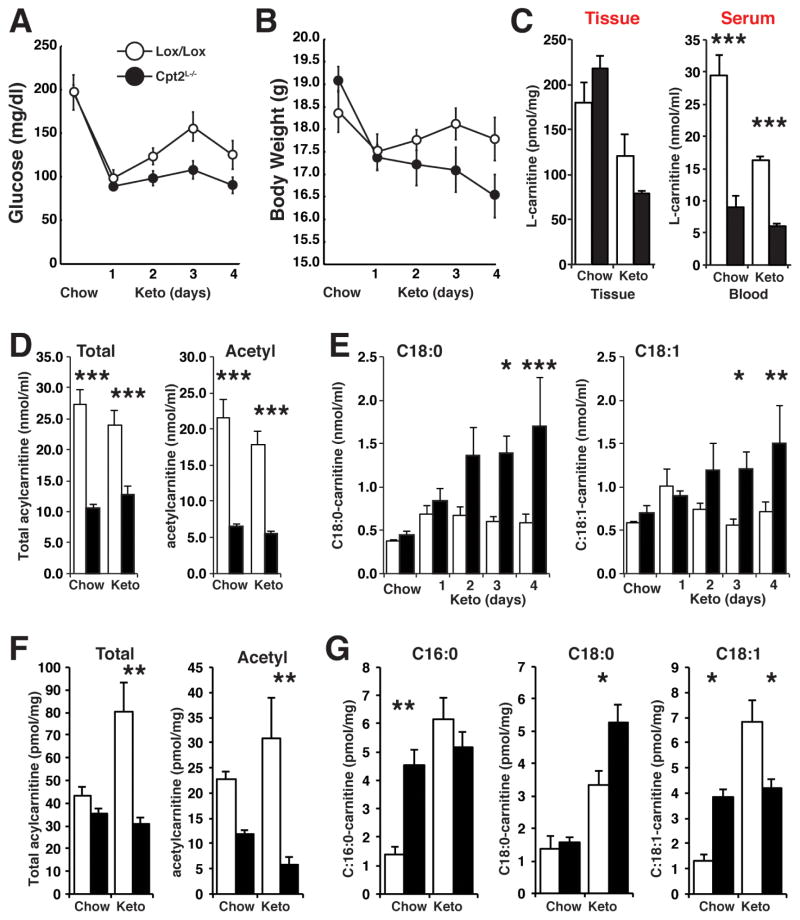
The liver is critical for maintaining systemic energy balance during starvation. To understand the role of hepatic fatty acid β-oxidation on this process, we generated mice with a liver-specific knockout of carnitine palmitoyltransferase 2 (Cpt2(L-/-)), an obligate step in mitochondrial long-chain fatty acid β-oxidation. Fasting induced hepatic steatosis and serum dyslipidemia with an absence of circulating ketones, while blood glucose remained normal. Systemic energy homeostasis was largely maintained in fasting Cpt2(L-/-) mice by adaptations in hepatic and systemic oxidative gene expression mediated in part by Pparα target genes including procatabolic hepatokines Fgf21, Gdf15, and Igfbp1. Feeding a ketogenic diet to Cpt2(L-/-) mice resulted in severe hepatomegaly, liver damage, and death with a complete absence of adipose triglyceride stores. These data show that hepatic fatty acid oxidation is not required for survival during acute food deprivation but essential for constraining adipocyte lipolysis and regulating systemic catabolism when glucose is limiting.
Copyright © 2016 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors have no competing financial interests.
Figures
References
-
- Cahill GF., Jr Fuel metabolism in starvation. Annu Rev Nutr. 2006;26:1–22. - PubMed
-
- Mutel E, Gautier-Stein A, Abdul-Wahed A, Amigo-Correig M, Zitoun C, Stefanutti A, Houberdon I, Tourette JA, Mithieux G, Rajas F. Control of blood glucose in the absence of hepatic glucose production during prolonged fasting in mice: induction of renal and intestinal gluconeogenesis by glucagon. Diabetes. 2011;60(12):3121–31. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
