

Neuronal activity enhances tau propagation and tau pathology in vivo
- PMID: 27322420
- PMCID: PMC4961585
- DOI: 10.1038/nn.4328
Neuronal activity enhances tau propagation and tau pathology in vivo
Abstract
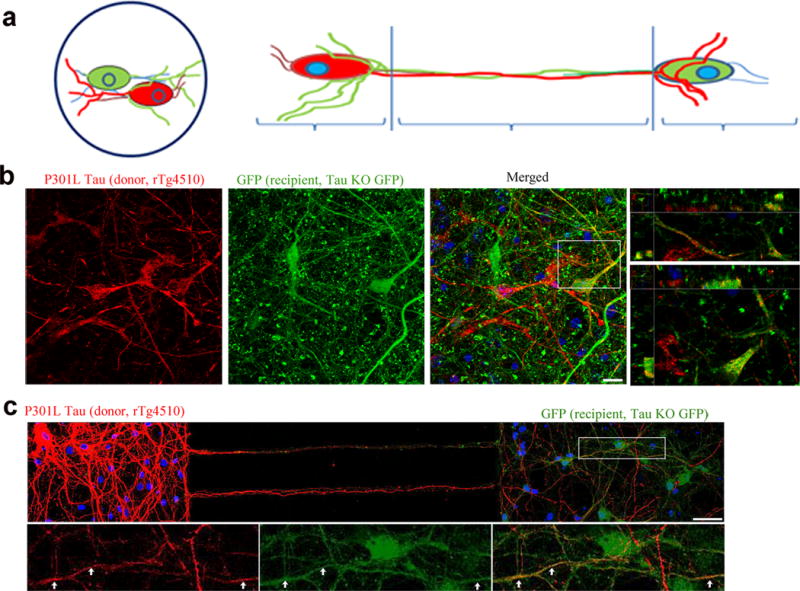
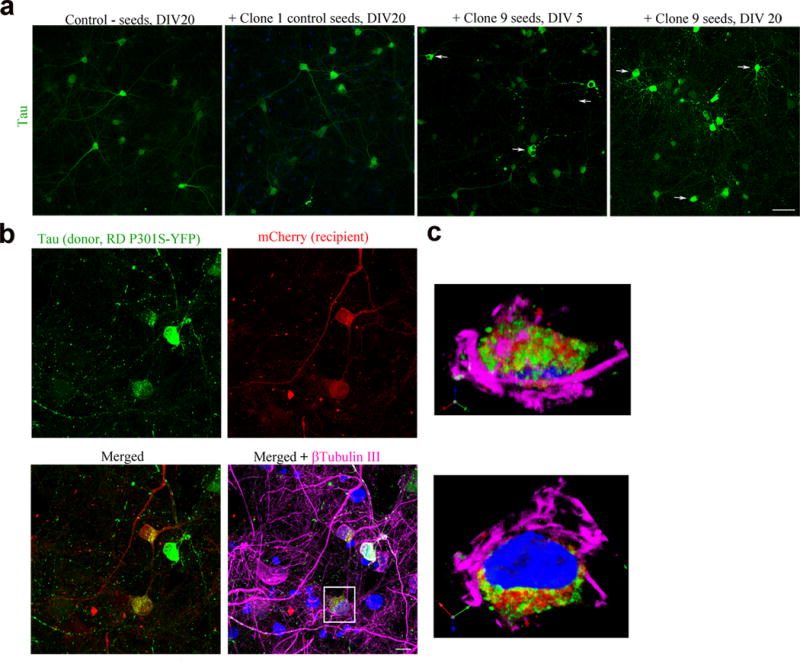
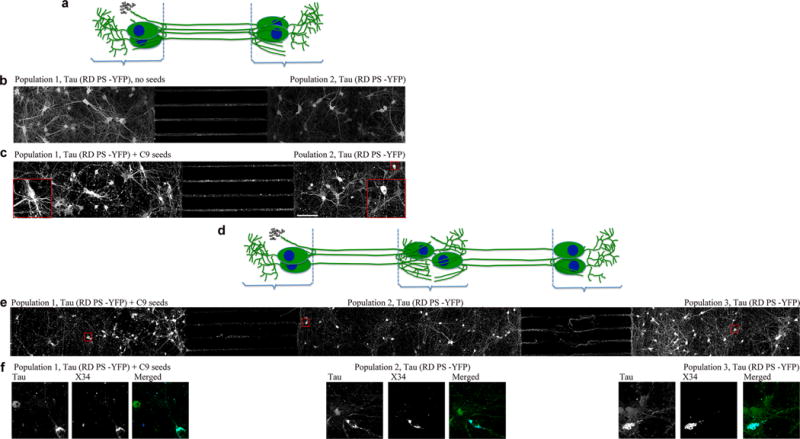
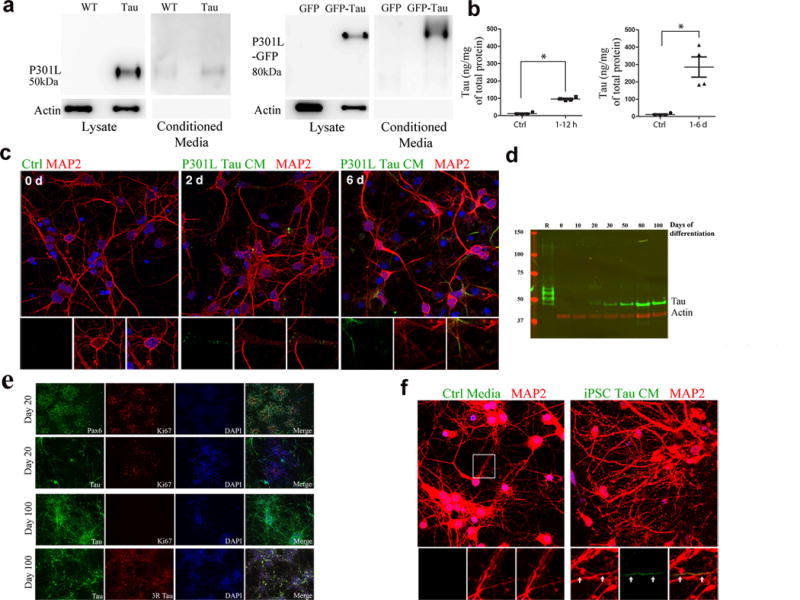
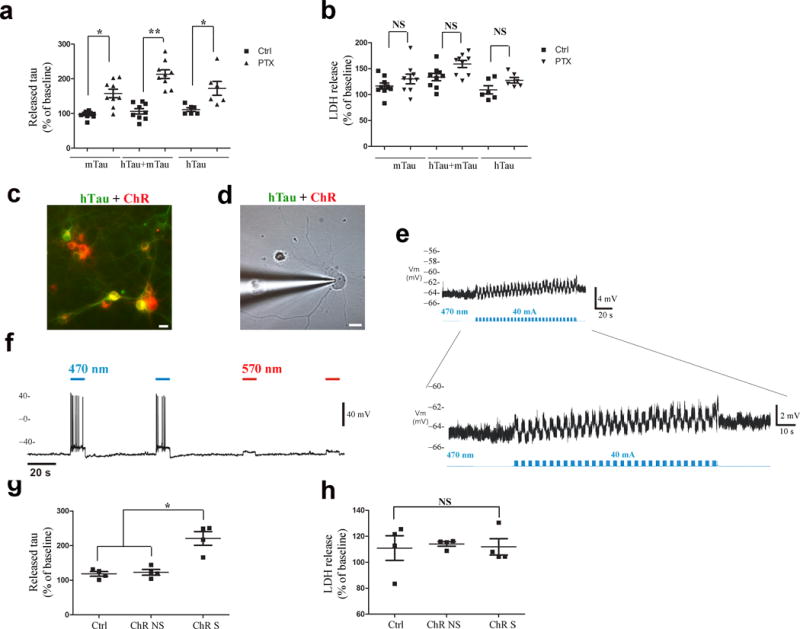
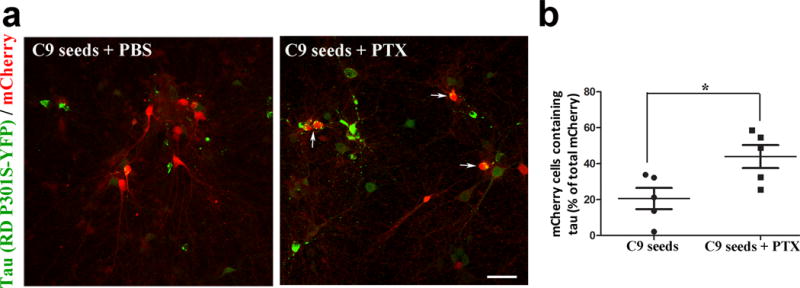
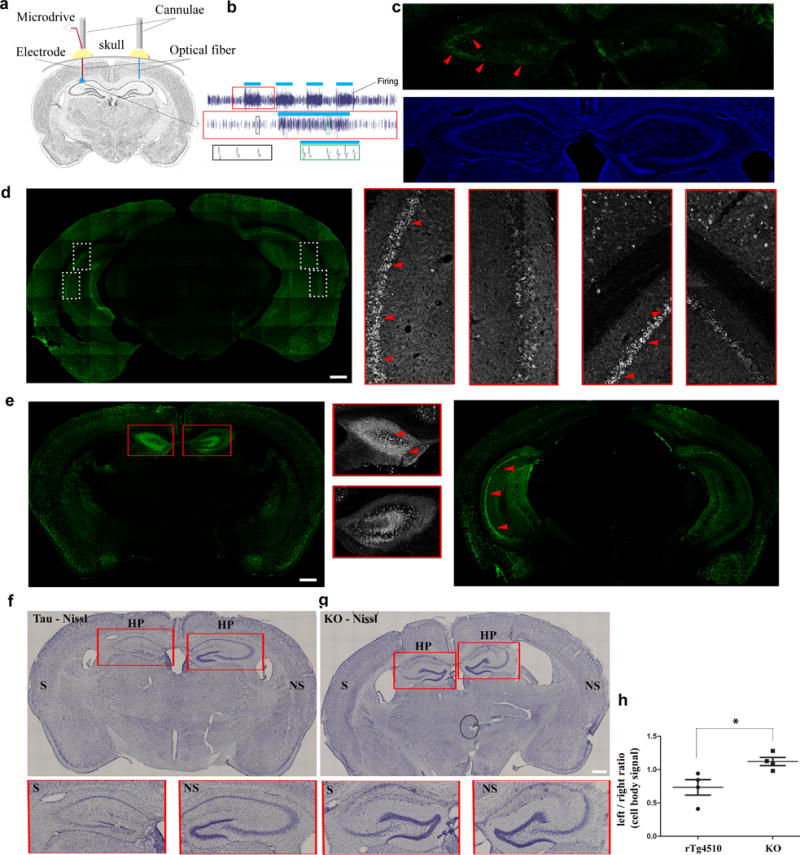
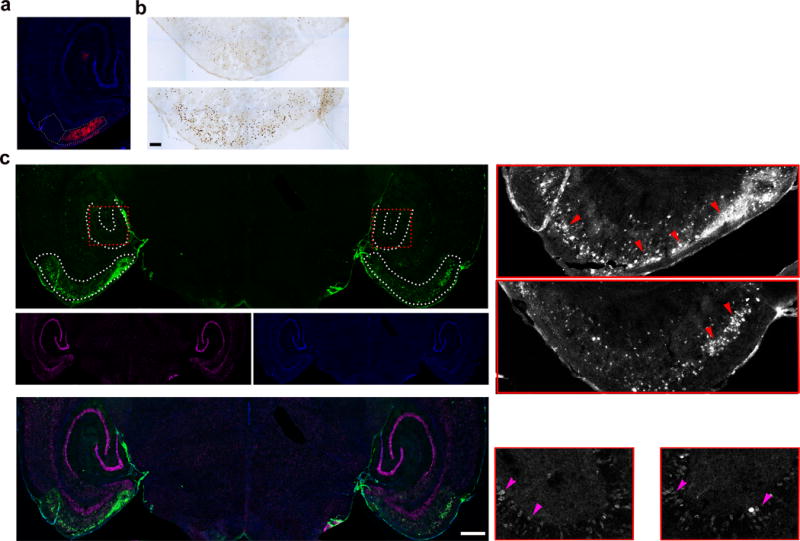
Tau protein can transfer between neurons transneuronally and trans-synaptically, which is thought to explain the progressive spread of tauopathy observed in the brain of patients with Alzheimer's disease. Here we show that physiological tau released from donor cells can transfer to recipient cells via the medium, suggesting that at least one mechanism by which tau can transfer is via the extracellular space. Neuronal activity has been shown to regulate tau secretion, but its effect on tau pathology is unknown. Using optogenetic and chemogenetic approaches, we found that increased neuronal activity stimulates the release of tau in vitro and enhances tau pathology in vivo. These data have implications for disease pathogenesis and therapeutic strategies for Alzheimer's disease and other tauopathies.
Conflict of interest statement
Competing Financial Interests
The authors declare no competing financial interests.
Figures
References
-
- Braak H, Braak E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991;82:239–259. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
