

Filopodial-Tension Model of Convergent-Extension of Tissues
- PMID: 27322528
- PMCID: PMC4913901
- DOI: 10.1371/journal.pcbi.1004952
Filopodial-Tension Model of Convergent-Extension of Tissues
Abstract
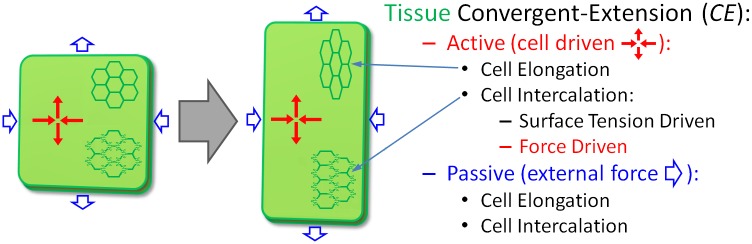
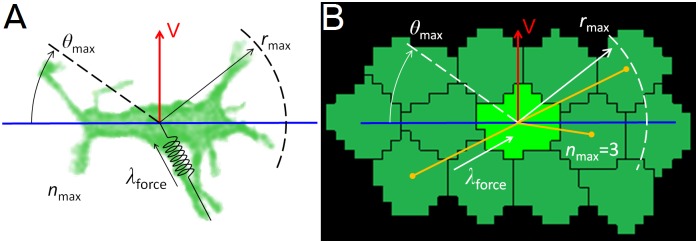
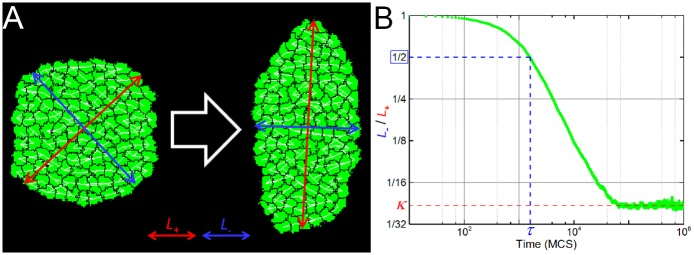
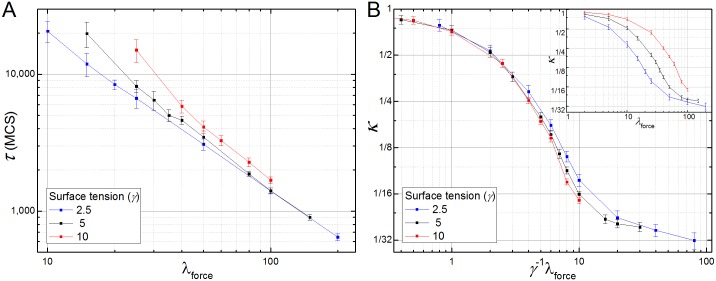
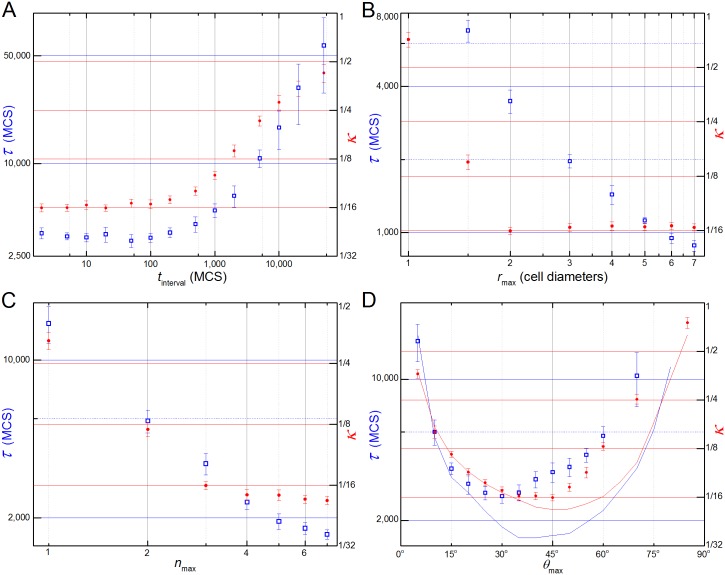
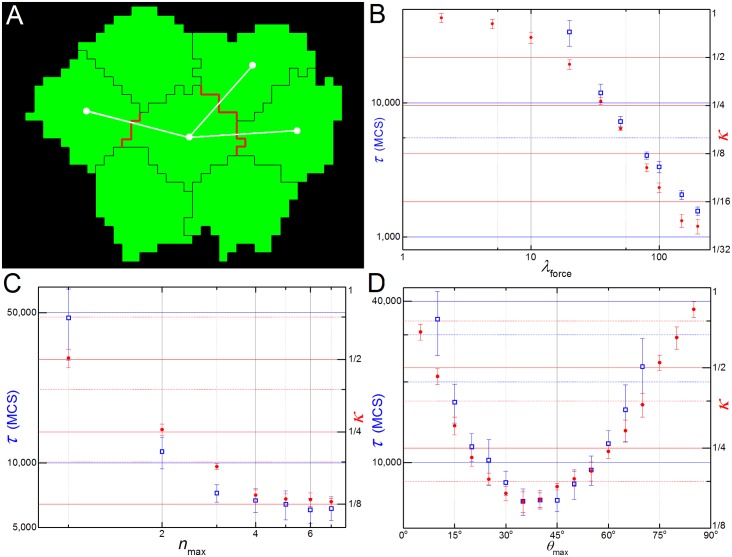
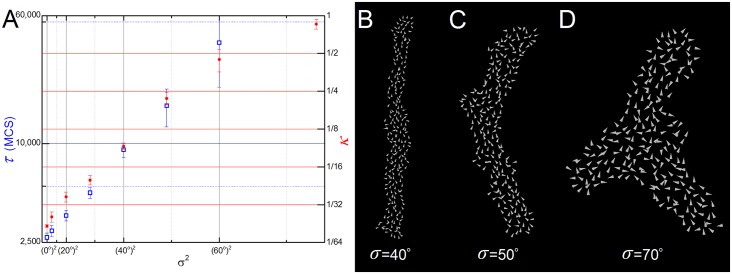
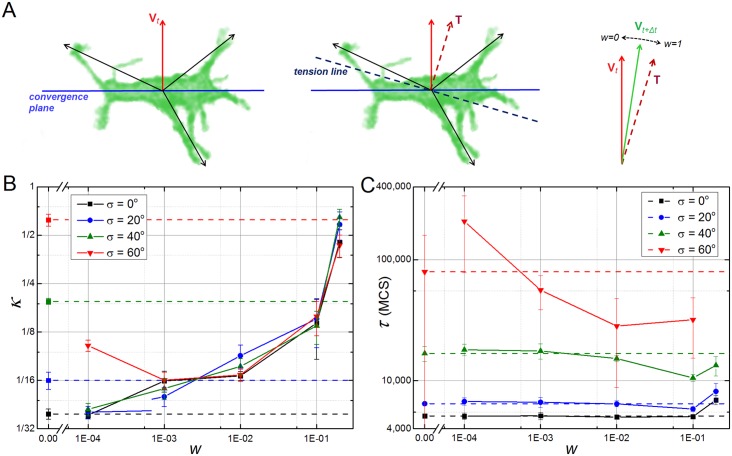
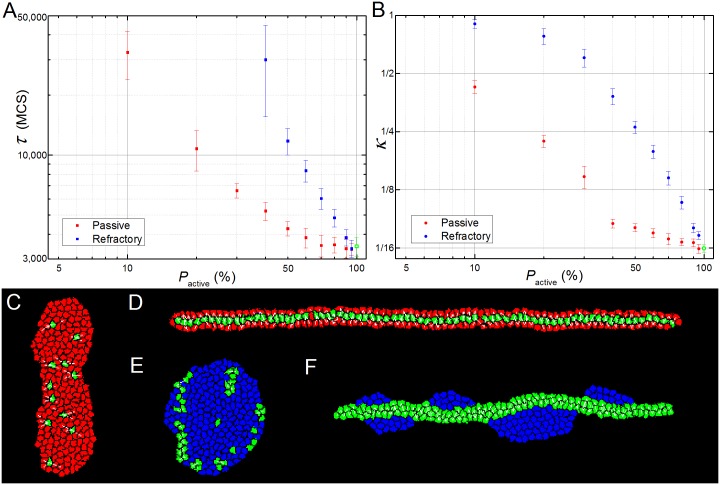
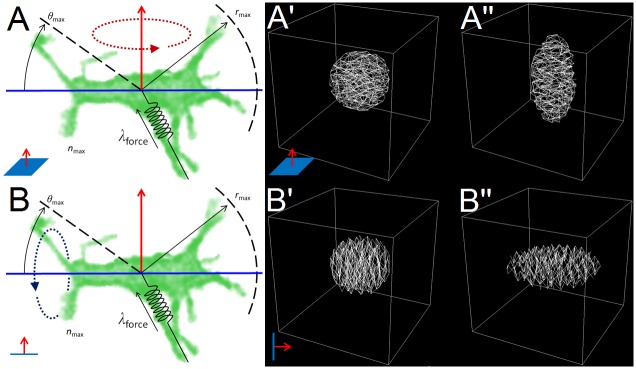
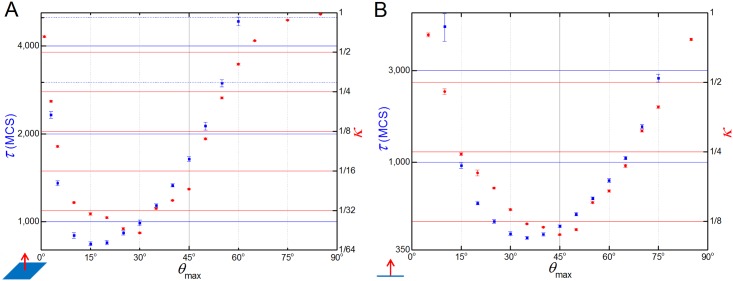
In convergent-extension (CE), a planar-polarized epithelial tissue elongates (extends) in-plane in one direction while shortening (converging) in the perpendicular in-plane direction, with the cells both elongating and intercalating along the converging axis. CE occurs during the development of most multicellular organisms. Current CE models assume cell or tissue asymmetry, but neglect the preferential filopodial activity along the convergent axis observed in many tissues. We propose a cell-based CE model based on asymmetric filopodial tension forces between cells and investigate how cell-level filopodial interactions drive tissue-level CE. The final tissue geometry depends on the balance between external rounding forces and cell-intercalation traction. Filopodial-tension CE is robust to relatively high levels of planar cell polarity misalignment and to the presence of non-active cells. Addition of a simple mechanical feedback between cells fully rescues and even improves CE of tissues with high levels of polarity misalignments. Our model extends easily to three dimensions, with either one converging and two extending axes, or two converging and one extending axes, producing distinct tissue morphologies, as observed in vivo.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Glickman NS, Kimmel CB, Jones MA, Adams RJ (2003) Shaping the zebrafish notochord. Development 130: 873–887. - PubMed
-
- Keller R, Tibbetts P (1989) Mediolateral Cell Intercalation in the Dorsal, Axial Mesoderm of Xenopus-Laevis. Developmental Biology 131: 539–549. - PubMed
-
- Davidson LA, Keller RE (1999) Neural tube closure in Xenopus laevis involves medial migration, directed protrusive activity, cell intercalation and convergent extension. Development 126: 4547–4556. - PubMed
-
- Voiculescu O, Bertocchini F, Wolpert L, Keller RE, Stern CD (2007) The amniote primitive streak is defined by epithelial cell intercalation before gastrulation. Nature 449: 1049–1052. - PubMed
-
- Ettensohn CA (1985) Gastrulation in the Sea-Urchin Embryo Is Accompanied by the Rearrangement of Invaginating Epithelial-Cells. Developmental Biology 112: 383–390. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
