

Mycolactone-mediated neurite degeneration and functional effects in cultured human and rat DRG neurons: Mechanisms underlying hypoalgesia in Buruli ulcer
- PMID: 27325560
- PMCID: PMC4956182
- DOI: 10.1177/1744806916654144
Mycolactone-mediated neurite degeneration and functional effects in cultured human and rat DRG neurons: Mechanisms underlying hypoalgesia in Buruli ulcer
Abstract
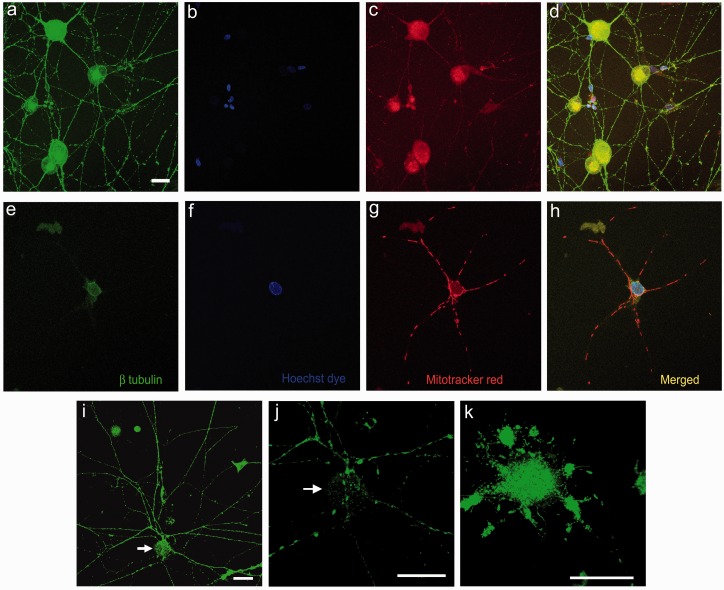
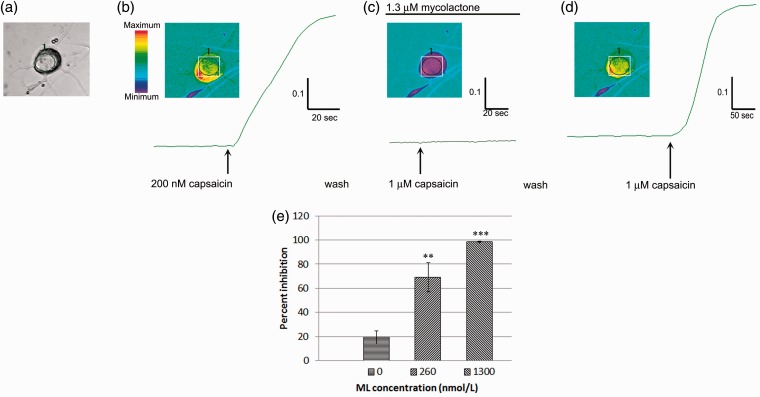
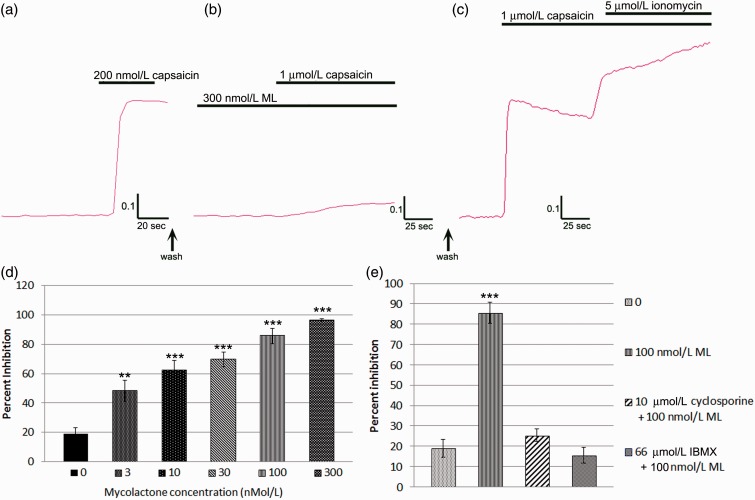
Background: Mycolactone is a polyketide toxin secreted by the mycobacterium Mycobacterium ulcerans, responsible for the extensive hypoalgesic skin lesions characteristic of patients with Buruli ulcer. A recent pre-clinical study proposed that mycolactone may produce analgesia via activation of the angiotensin II type 2 receptor (AT2R). In contrast, AT2R antagonist EMA401 has shown analgesic efficacy in animal models and clinical trials for neuropathic pain. We therefore investigated the morphological and functional effects of mycolactone in cultured human and rat dorsal root ganglia (DRG) neurons and the role of AT2R using EMA401. Primary sensory neurons were prepared from avulsed cervical human DRG and rat DRG; 24 h after plating, neurons were incubated for 24 to 96 h with synthetic mycolactone A/B, followed by immunostaining with antibodies to PGP9.5, Gap43, β tubulin, or Mitotracker dye staining. Acute functional effects were examined by measuring capsaicin responses with calcium imaging in DRG neuronal cultures treated with mycolactone.
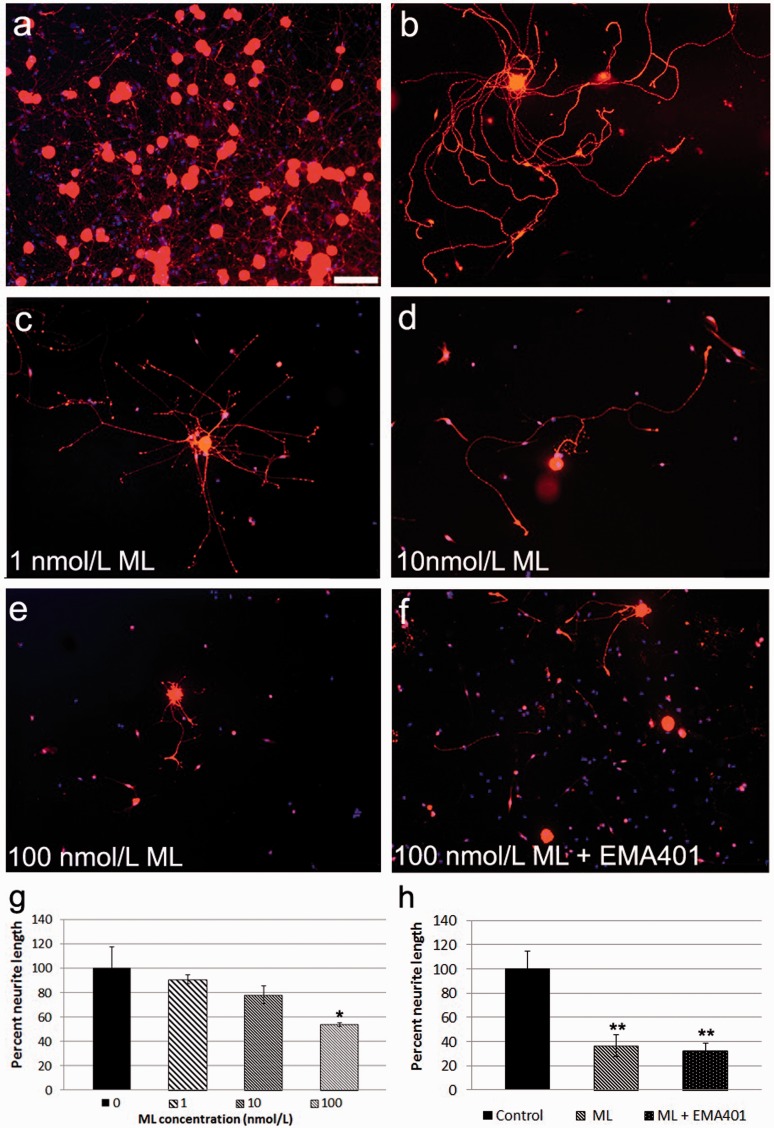
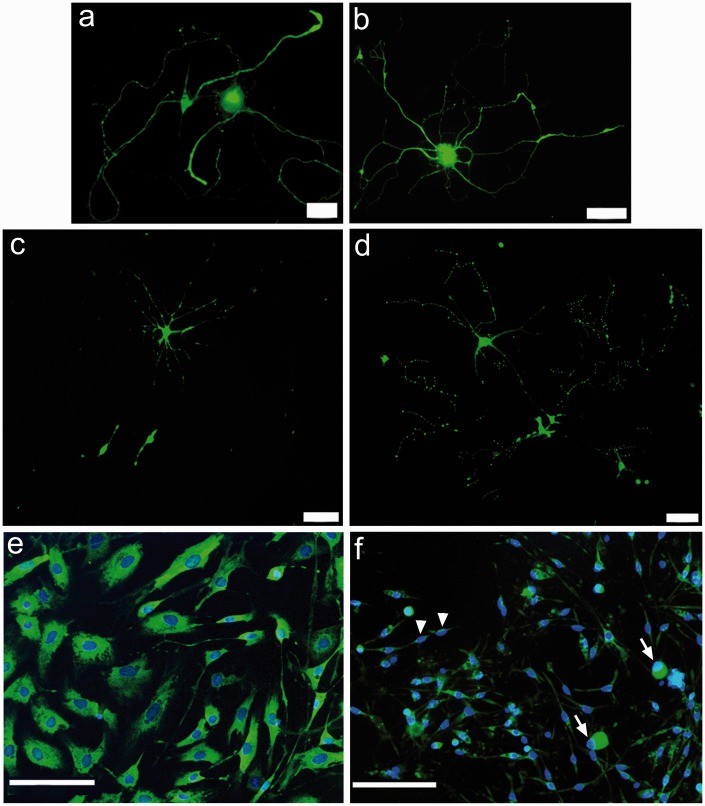
Results: Morphological effects: Mycolactone-treated cultures showed dramatically reduced numbers of surviving neurons and non-neuronal cells, reduced Gap43 and β tubulin expression, degenerating neurites and reduced cell body diameter, compared with controls. Dose-related reduction of neurite length was observed in mycolactone-treated cultures. Mitochondria were distributed throughout the length of neurites and soma of control neurons, but clustered in the neurites and soma of mycolactone-treated neurons. Functional effects: Mycolactone-treated human and rat DRG neurons showed dose-related inhibition of capsaicin responses, which were reversed by calcineurin inhibitor cyclosporine and phosphodiesterase inhibitor 3-isobutyl-1-Methylxanthine, indicating involvement of cAMP/ATP reduction. The morphological and functional effects of mycolactone were not altered by Angiotensin II or AT2R antagonist EMA401.
Conclusion: Mycolactone induces toxic effects in DRG neurons, leading to impaired nociceptor function, neurite degeneration, and cell death, resembling the cutaneous hypoalgesia and nerve damage in individuals with M. Ulcerans infection.
Keywords: Buruli ulcer; TRPV1; apoptosis; calcium influx; hypoalgesia; mitochondria; mycolactone; neurite degeneration; neurons.
© The Author(s) 2016.
Figures
References
-
- WHO. Treatment of Mycobacterium Ulcerans Disease (Buruli Ulcer). Guidance for Health Workers, 2012; www.who.int/mediacentre/factsheets/fs199/en/ (accessed 1 June 2016).
-
- Einarsdottir T, Huygen K. Buruli ulcer. Hum Vaccines 2011; 7: 1198–1203. - PubMed
-
- Walsh DS, Prttaels F, Myers WM. Buruli ulcer: advances in understanding Mycobacterium Ulcerans infection. Dermatol Clin 2011; 29: 1–8. - PubMed
-
- Bozzo C, Tiberio R, Graziola F, et al. A Mycobacterium ulcerans toxin, mycolactone, induces apoptosis in primary human keratinocytes and in HaCaT cells. Microbes Infect 2010; 12: 1258–1263. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
