

Pancreatic β-Cell production of CXCR3 ligands precedes diabetes onset
- PMID: 27325565
- PMCID: PMC5177512
- DOI: 10.1002/biof.1304
Pancreatic β-Cell production of CXCR3 ligands precedes diabetes onset
Abstract
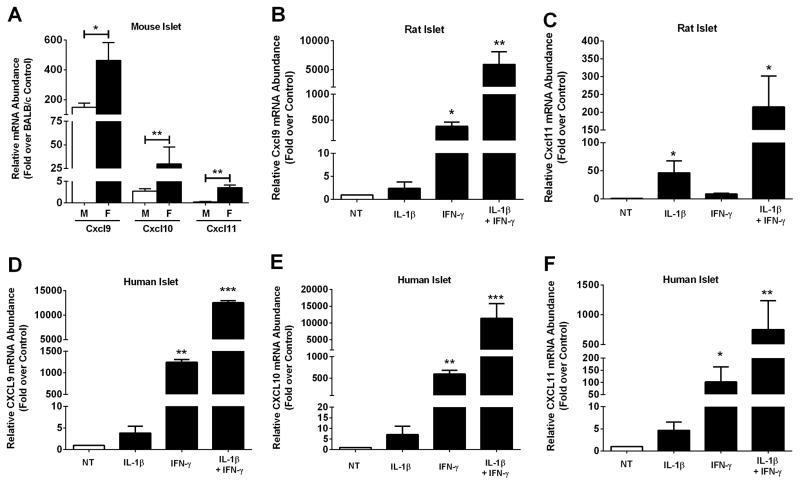
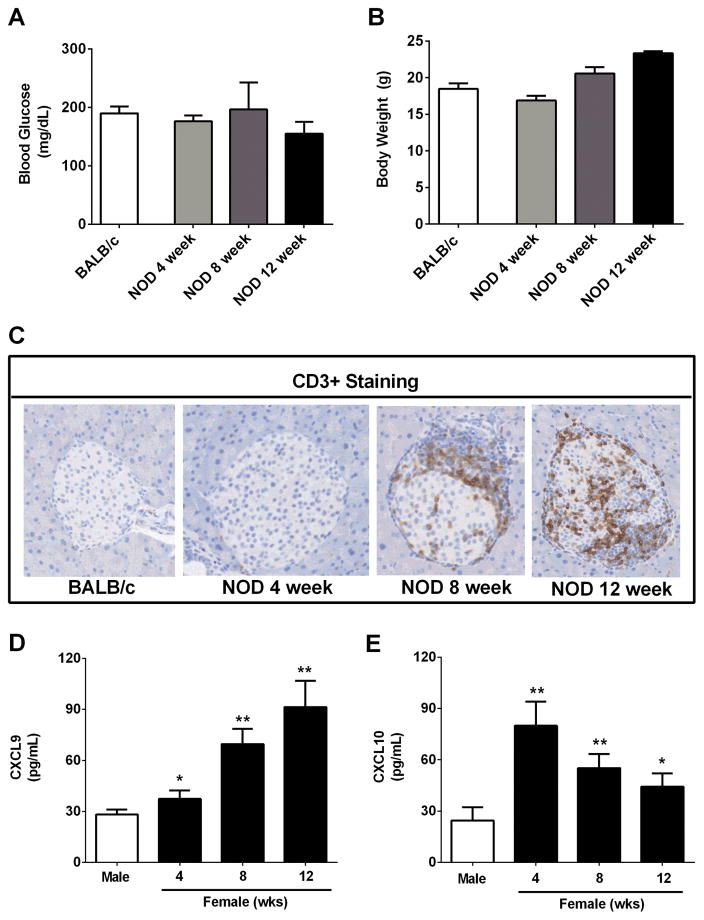
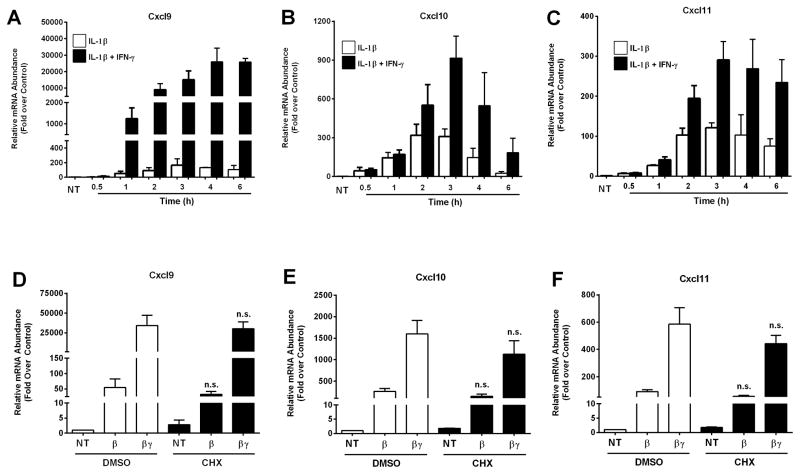
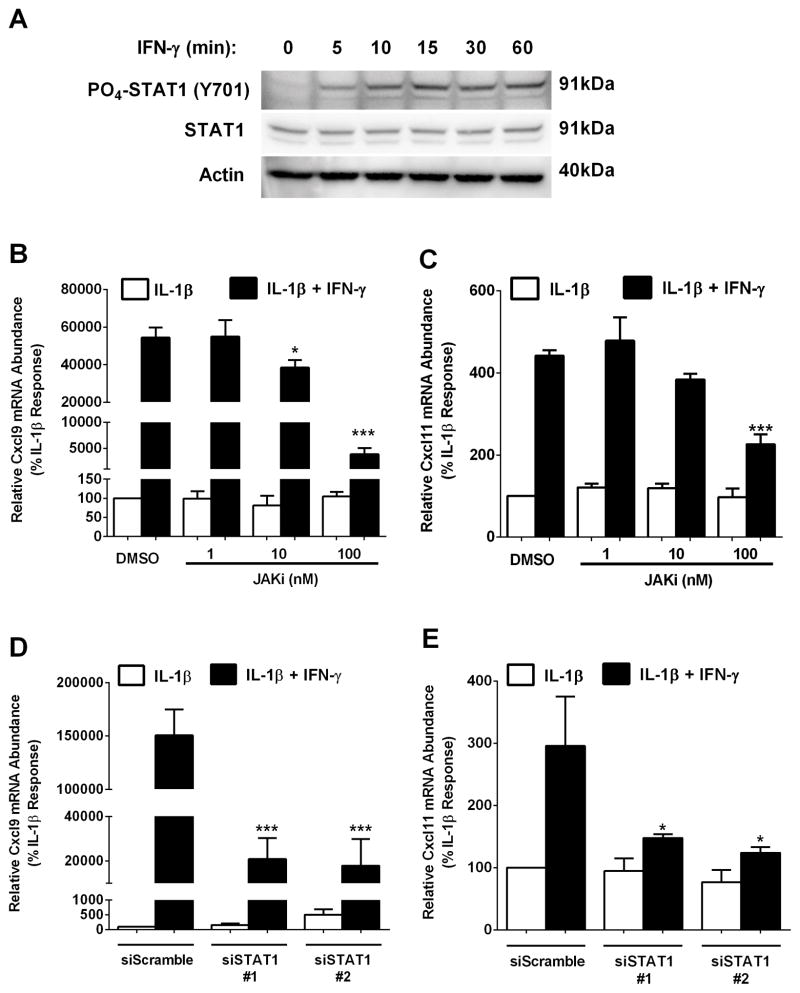
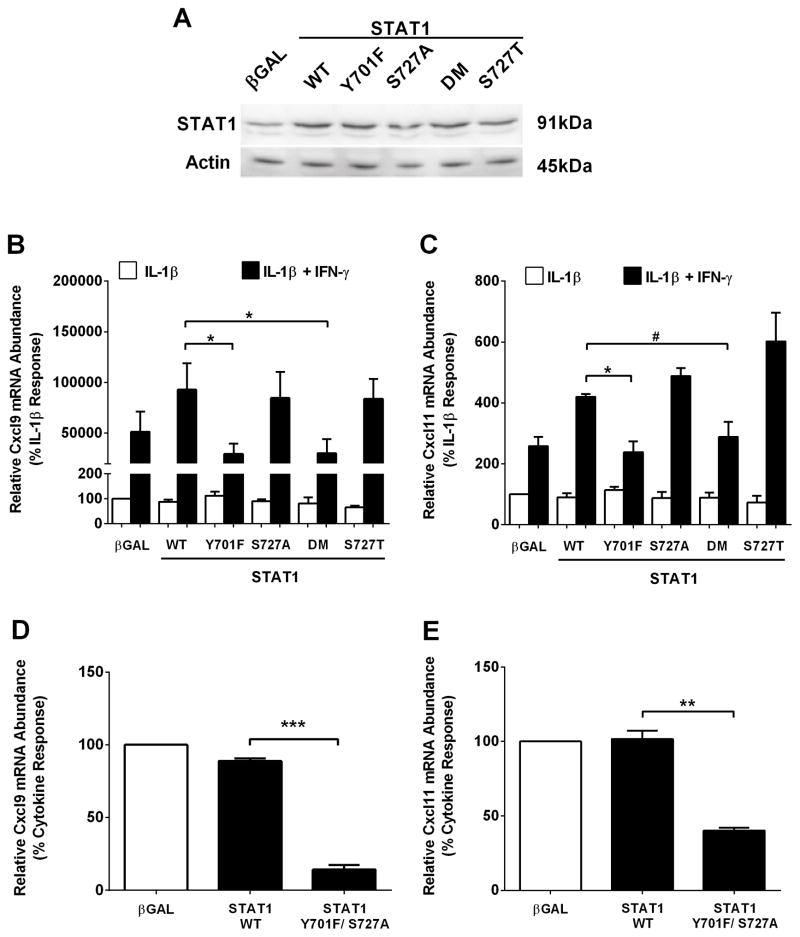
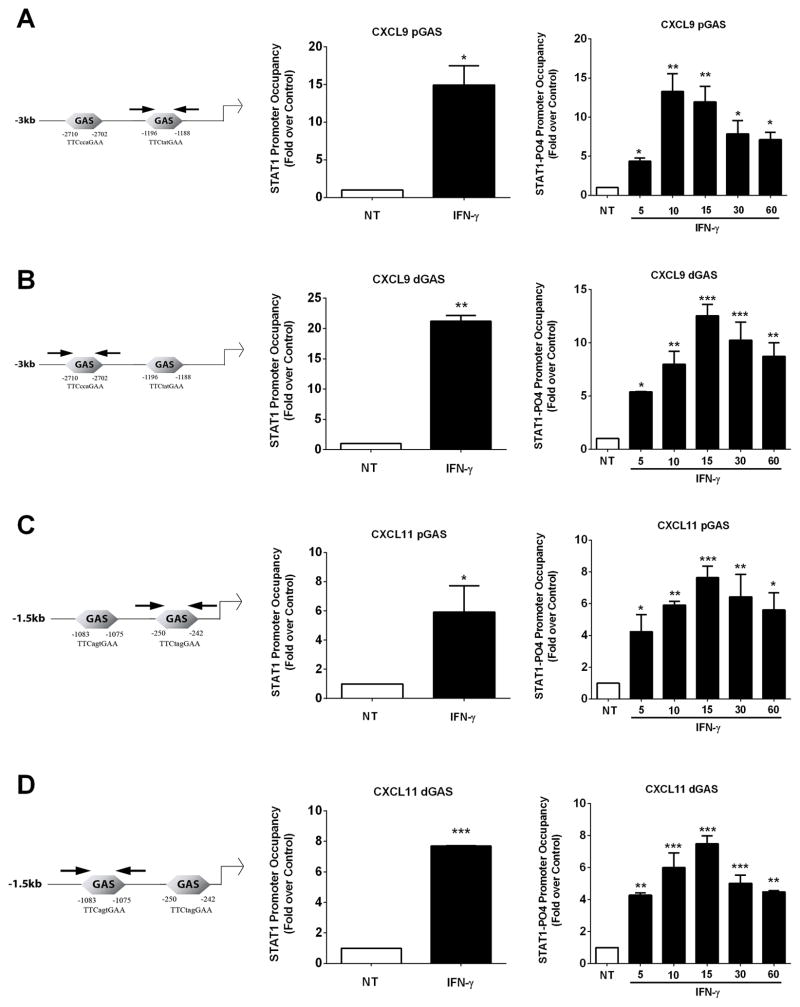
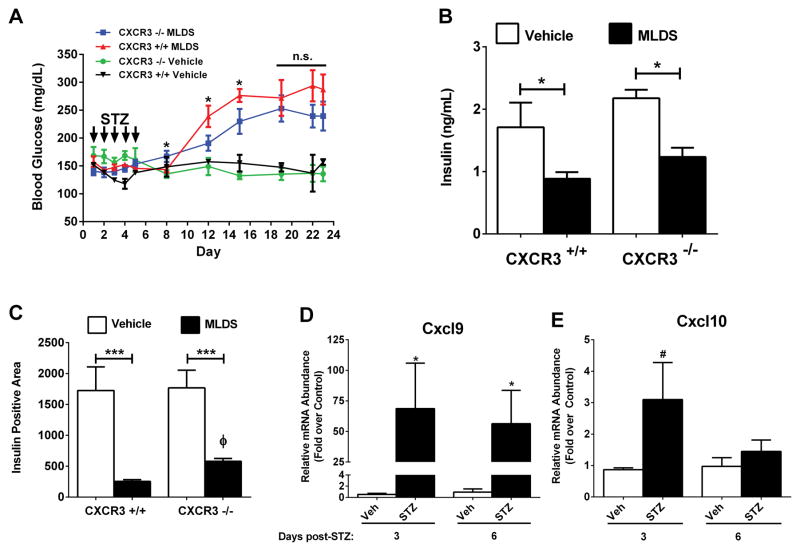
Type 1 diabetes mellitus (T1DM) results from immune cell-mediated reductions in function and mass of the insulin-producing β-cells within the pancreatic islets. While the initial trigger(s) that initiates the autoimmune process is unknown, there is a leukocytic infiltration that precedes islet β-cell death and dysfunction. Herein, we demonstrate that genes encoding the chemokines CXCL9, 10, and 11 are primary response genes in pancreatic β-cells and are also elevated as part of the inflammatory response in mouse, rat, and human islets. We further established that STAT1 participates in the transcriptional control of these genes in response to the pro-inflammatory cytokines IL-1β and IFN-γ. STAT1 is phosphorylated within five minutes after β-cell exposure to IFN-γ, with subsequent occupancy at proximal and distal response elements within the Cxcl9 and Cxcl11 gene promoters. This increase in STAT1 binding is coupled to the rapid appearance of chemokine transcript. Moreover, circulating levels of chemokines that activate CXCR3 are elevated in non-obese diabetic (NOD) mice, consistent with clinical findings in human diabetes. We also report herein that mice with genetic deletion of CXCR3 (receptor for ligands CXCL9, 10, and 11) exhibit a delay in diabetes development after being injected with multiple low doses of streptozotocin. Therefore, we conclude that production of CXCL9, 10, and 11 from islet β-cells controls leukocyte migration and activity into pancreatic tissue, which ultimately influences islet β-cell mass and function. © 2016 BioFactors, 42(6):703-715, 2016.
Keywords: autoimmunity; chemokine; diabetes; inflammation; islets; transcription.
© 2016 International Union of Biochemistry and Molecular Biology.
Conflict of interest statement
The authors have no conflicts of interest to disclose.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
