

Light Remodels Lipid Biosynthesis in Nannochloropsis gaditana by Modulating Carbon Partitioning between Organelles
- PMID: 27325666
- PMCID: PMC4972283
- DOI: 10.1104/pp.16.00599
Light Remodels Lipid Biosynthesis in Nannochloropsis gaditana by Modulating Carbon Partitioning between Organelles
Abstract
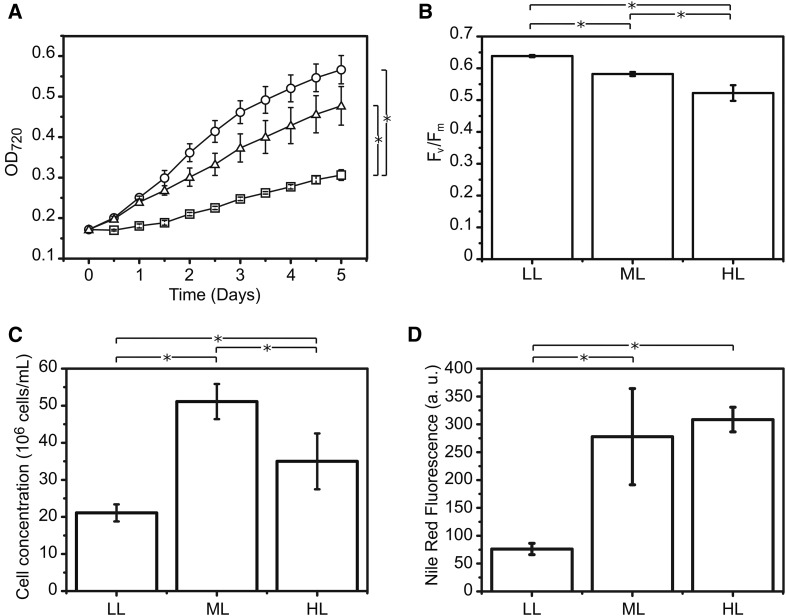
The seawater microalga Nannochloropsis gaditana is capable of accumulating a large fraction of reduced carbon as lipids. To clarify the molecular bases of this metabolic feature, we investigated light-driven lipid biosynthesis in Nannochloropsis gaditana cultures combining the analysis of photosynthetic functionality with transcriptomic, lipidomic and metabolomic approaches. Light-dependent alterations are observed in amino acid, isoprenoid, nucleic acid, and vitamin biosynthesis, suggesting a deep remodeling in the microalgal metabolism triggered by photoadaptation. In particular, high light intensity is shown to affect lipid biosynthesis, inducing the accumulation of diacylglyceryl-N,N,N-trimethylhomo-Ser and triacylglycerols, together with the up-regulation of genes involved in their biosynthesis. Chloroplast polar lipids are instead decreased. This situation correlates with the induction of genes coding for a putative cytosolic fatty acid synthase of type 1 (FAS1) and polyketide synthase (PKS) and the down-regulation of the chloroplast fatty acid synthase of type 2 (FAS2). Lipid accumulation is accompanied by the regulation of triose phosphate/inorganic phosphate transport across the chloroplast membranes, tuning the carbon metabolic allocation between cell compartments, favoring the cytoplasm, mitochondrion, and endoplasmic reticulum at the expense of the chloroplast. These results highlight the high flexibility of lipid biosynthesis in N. gaditana and lay the foundations for a hypothetical mechanism of regulation of primary carbon partitioning by controlling metabolite allocation at the subcellular level.
© 2016 American Society of Plant Biologists. All Rights Reserved.
Figures







References
-
- Arrivault S, Guenther M, Ivakov A, Feil R, Vosloh D, van Dongen JT, Sulpice R, Stitt M (2009) Use of reverse-phase liquid chromatography, linked to tandem mass spectrometry, to profile the Calvin cycle and other metabolic intermediates in Arabidopsis rosettes at different carbon dioxide concentrations. Plant J 59: 826–839 - PubMed
-
- Bailey S, Walters RG, Jansson S, Horton P (2001) Acclimation of Arabidopsis thaliana to the light environment: the existence of separate low light and high light responses. Planta 213: 794–801 - PubMed
-
- Beer A, Gundermann K, Beckmann J, Büchel C (2006) Subunit composition and pigmentation of fucoxanthin-chlorophyll proteins in diatoms: evidence for a subunit involved in diadinoxanthin and diatoxanthin binding. Biochemistry 45: 13046–13053 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
