

The role of MscL amphipathic N terminus indicates a blueprint for bilayer-mediated gating of mechanosensitive channels
- PMID: 27329693
- PMCID: PMC4917966
- DOI: 10.1038/ncomms11984
The role of MscL amphipathic N terminus indicates a blueprint for bilayer-mediated gating of mechanosensitive channels
Abstract
The bacterial mechanosensitive channel MscL gates in response to membrane tension as a result of mechanical force transmitted directly to the channel from the lipid bilayer. MscL represents an excellent model system to study the basic biophysical principles of mechanosensory transduction. However, understanding of the essential structural components that transduce bilayer tension into channel gating remains incomplete. Here using multiple experimental and computational approaches, we demonstrate that the amphipathic N-terminal helix of MscL acts as a crucial structural element during tension-induced gating, both stabilizing the closed state and coupling the channel to the membrane. We propose that this may also represent a common principle in the gating cycle of unrelated mechanosensitive ion channels, allowing the coupling of channel conformation to membrane dynamics.
Figures
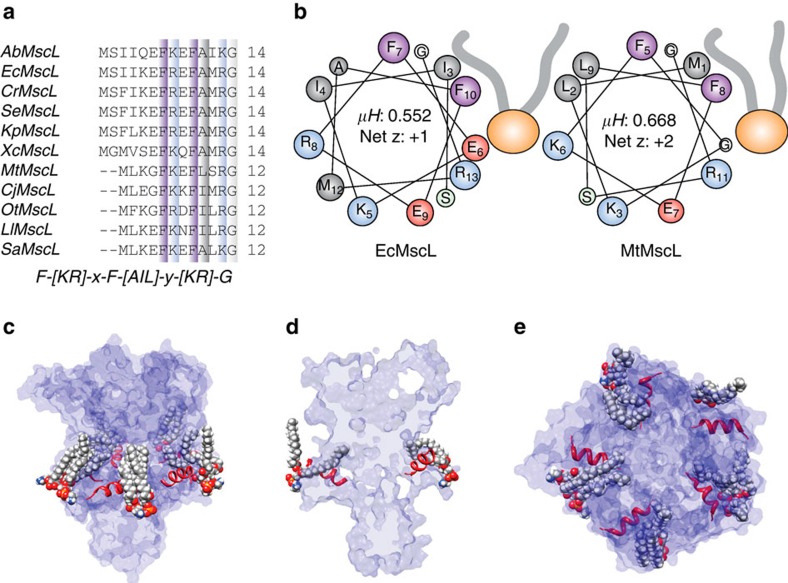
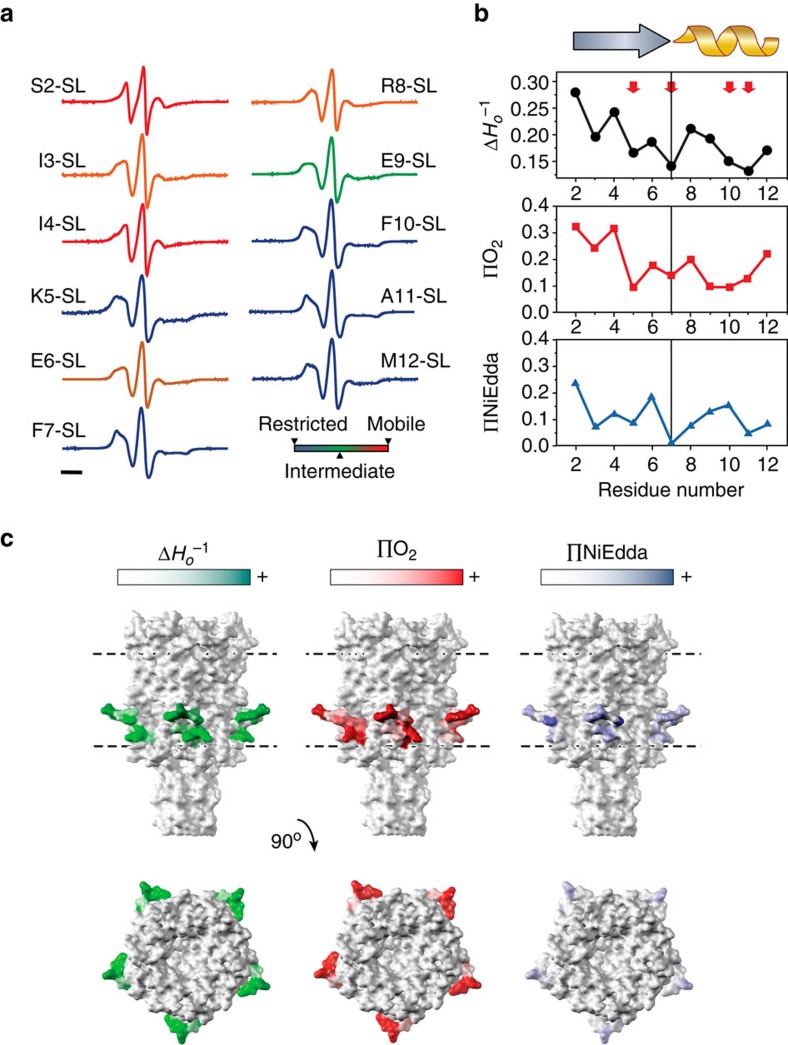
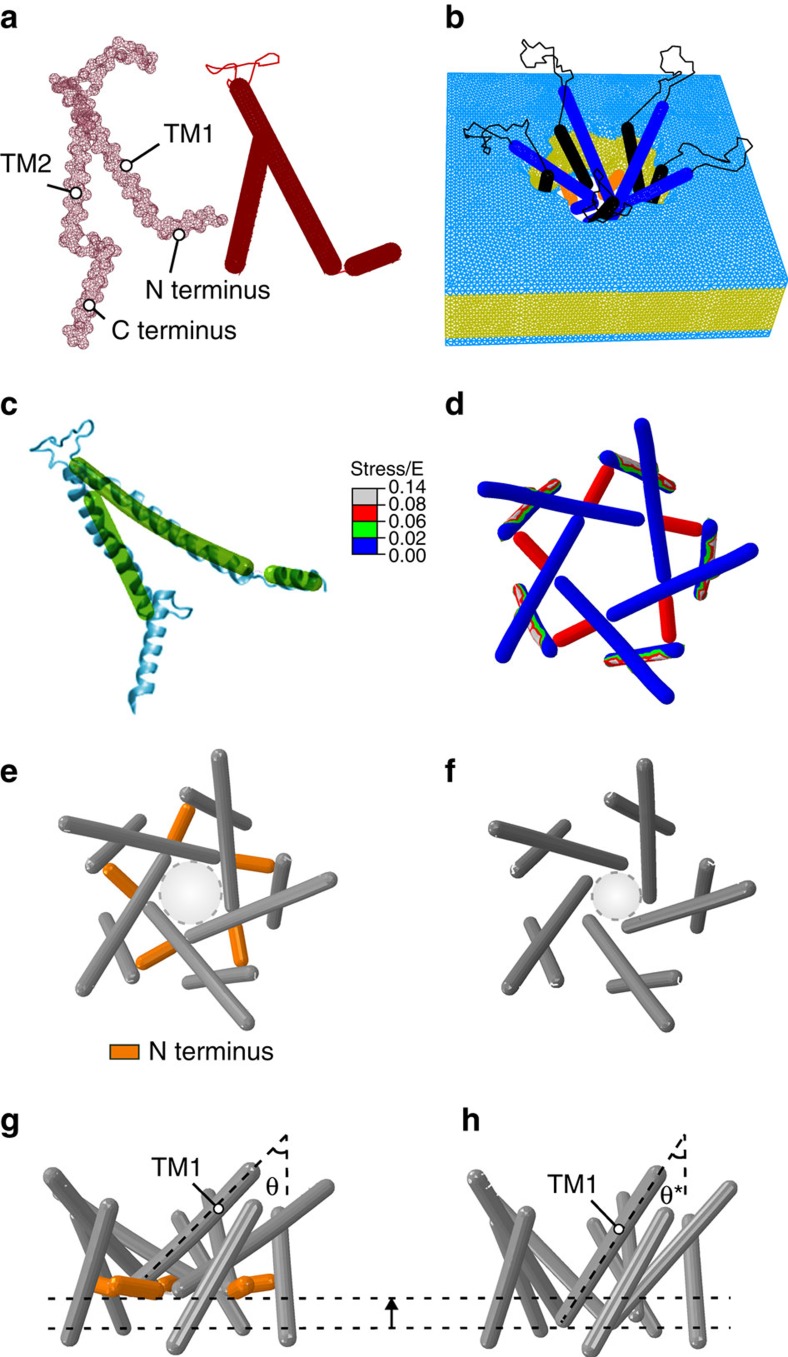
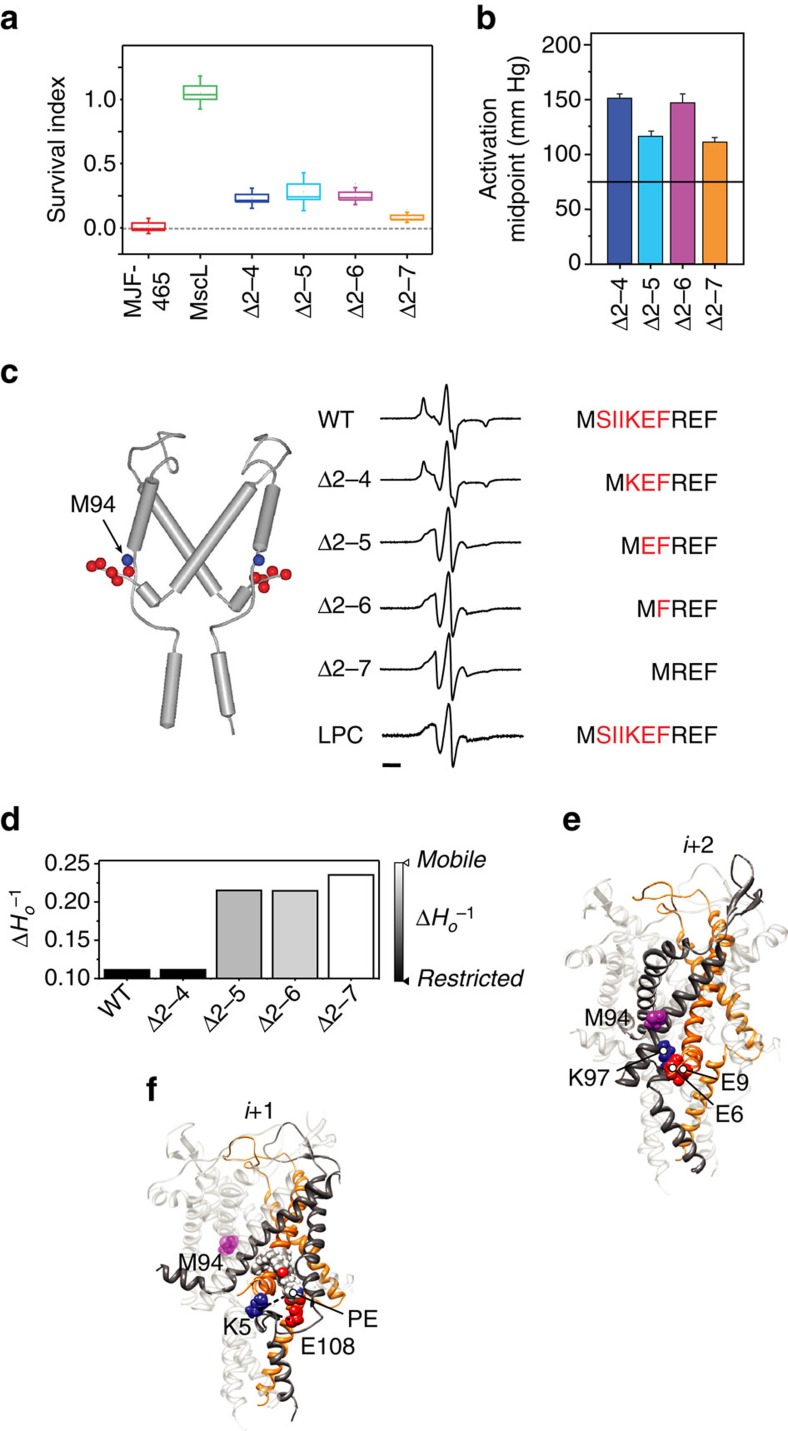
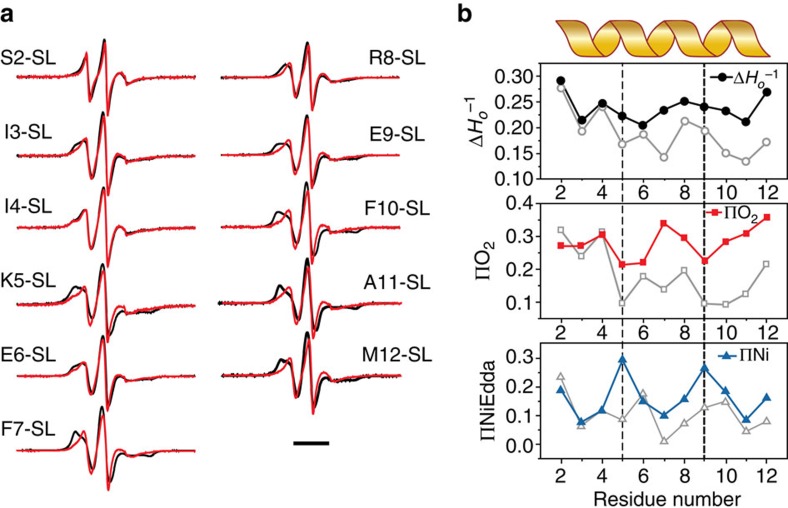
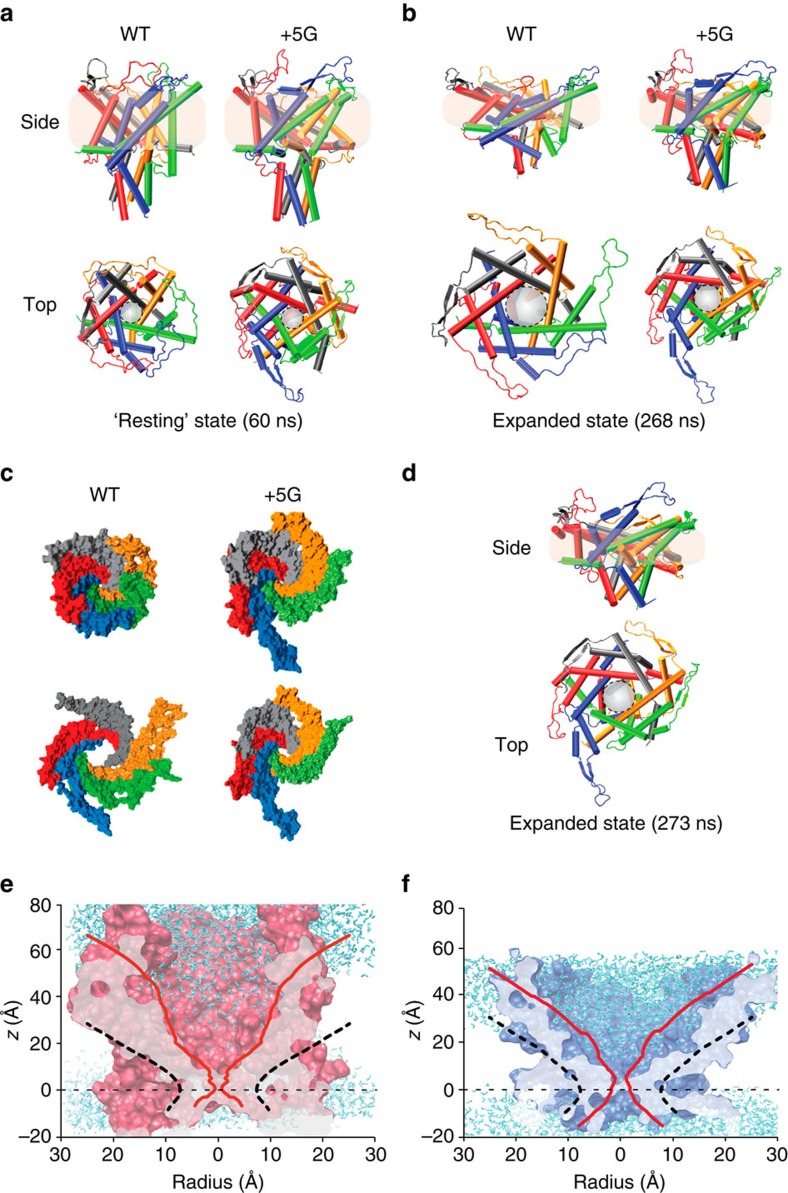
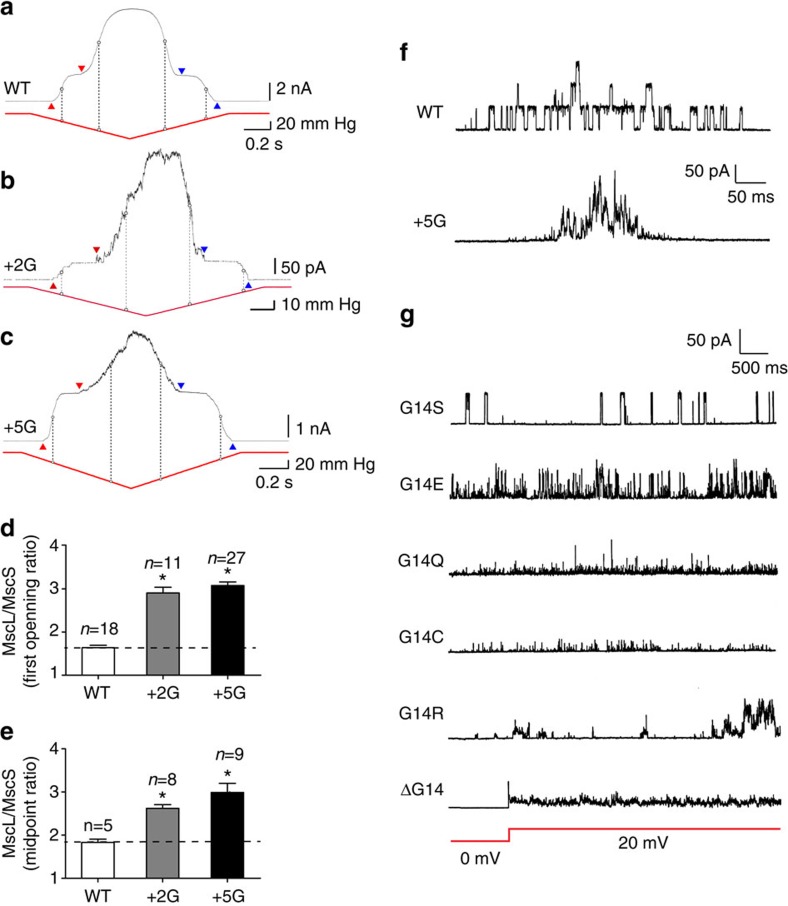
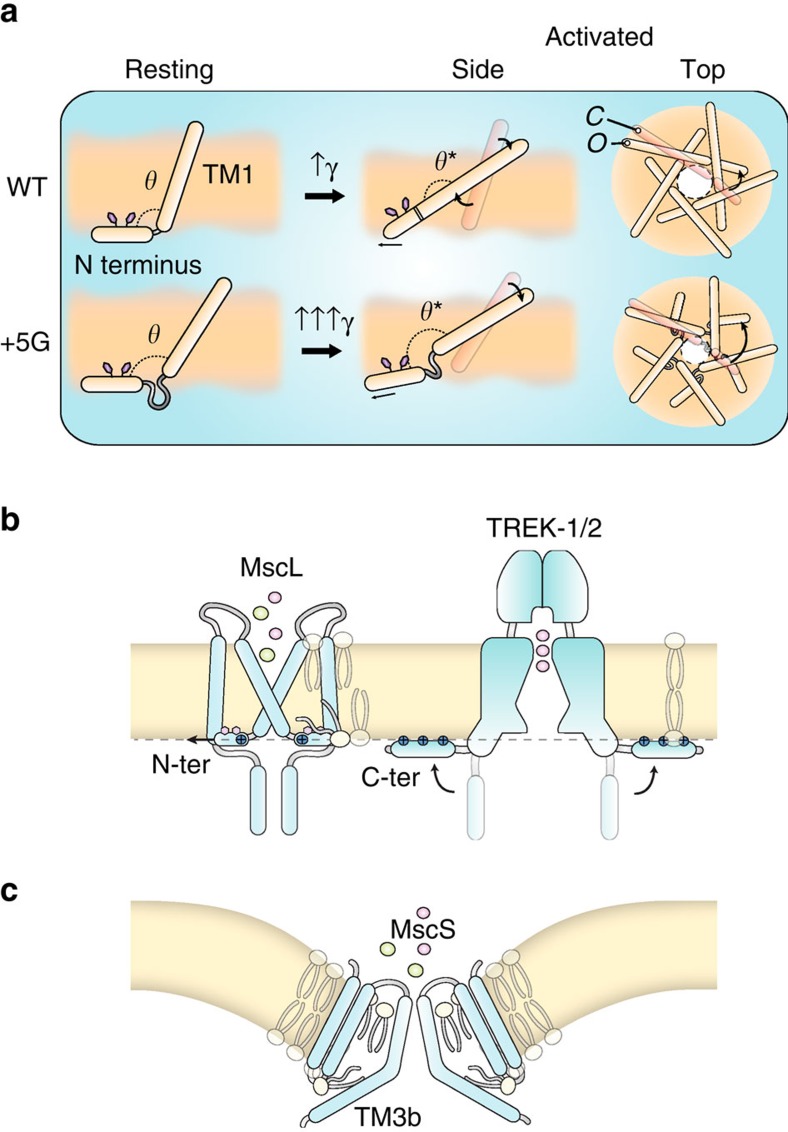
Comment in
-
Toward a structural blueprint for bilayer-mediated channel mechanosensitivity.Channels (Austin). 2017 Mar 4;11(2):91-93. doi: 10.1080/19336950.2016.1224624. Epub 2016 Aug 17. Channels (Austin). 2017. PMID: 27533603 Free PMC article. No abstract available.
References
-
- Hamill O. P. & Martinac B. Molecular basis of mechanotransduction in living cells. Physiol. Rev. 81, 685–740 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
