

Lipid signaling and lipotoxicity in metaflammation: indications for metabolic disease pathogenesis and treatment
- PMID: 27330055
- PMCID: PMC5321214
- DOI: 10.1194/jlr.R066514
Lipid signaling and lipotoxicity in metaflammation: indications for metabolic disease pathogenesis and treatment
Abstract
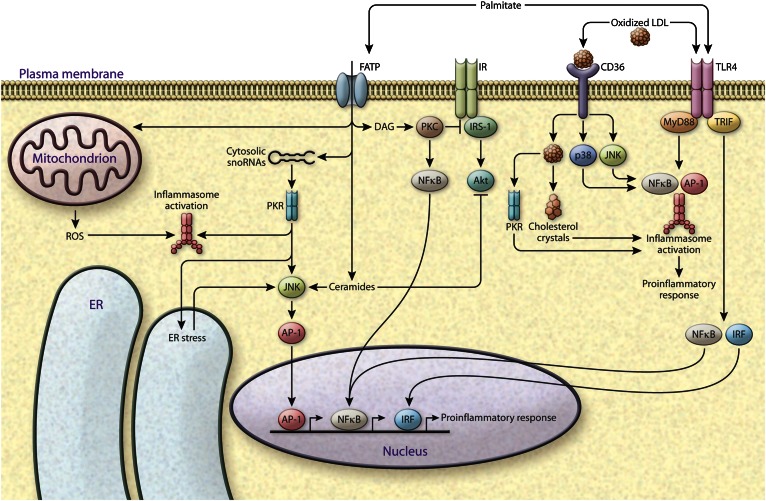
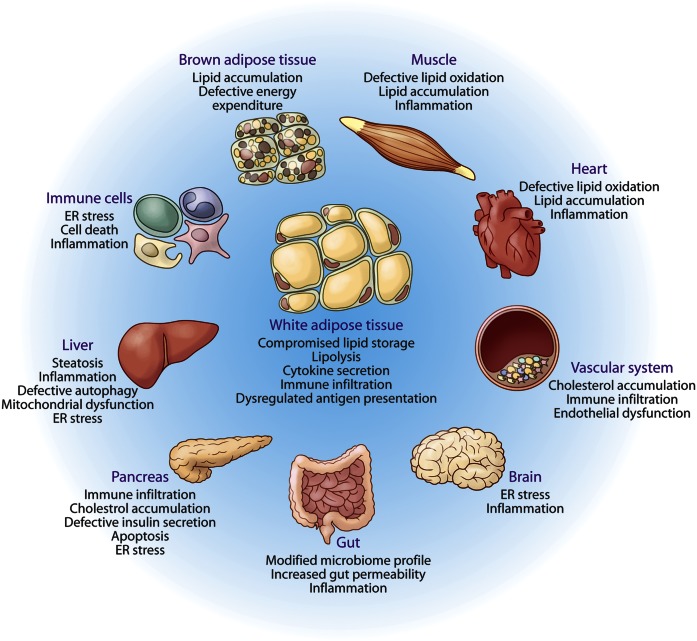
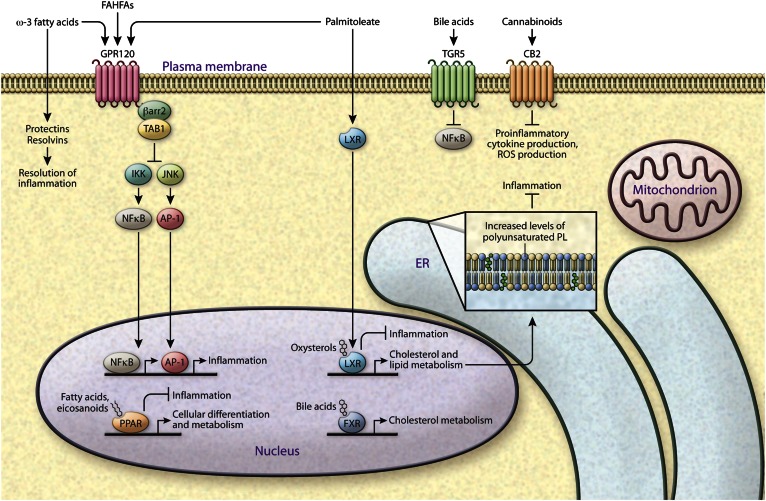
Lipids encompass a wide variety of molecules such as fatty acids, sterols, phospholipids, and triglycerides. These molecules represent a highly efficient energy resource and can act as structural elements of membranes or as signaling molecules that regulate metabolic homeostasis through many mechanisms. Cells possess an integrated set of response systems to adapt to stresses such as those imposed by nutrient fluctuations during feeding-fasting cycles. While lipids are pivotal for these homeostatic processes, they can also contribute to detrimental metabolic outcomes. When metabolic stress becomes chronic and adaptive mechanisms are overwhelmed, as occurs during prolonged nutrient excess or obesity, lipid influx can exceed the adipose tissue storage capacity and result in accumulation of harmful lipid species at ectopic sites such as liver and muscle. As lipid metabolism and immune responses are highly integrated, accumulation of harmful lipids or generation of signaling intermediates can interfere with immune regulation in multiple tissues, causing a vicious cycle of immune-metabolic dysregulation. In this review, we summarize the role of lipotoxicity in metaflammation at the molecular and tissue level, describe the significance of anti-inflammatory lipids in metabolic homeostasis, and discuss the potential of therapeutic approaches targeting pathways at the intersection of lipid metabolism and immune function.
Keywords: diabetes; inflammation; insulin resistance; lipids; obesity; signaling lipids.
Copyright © 2016 by the American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Hotamisligil G. S. 2006. Inflammation and metabolic disorders. Nature. 444: 860–867. - PubMed
-
- Fu S., Watkins S. M., and Hotamisligil G. S.. 2012. The role of endoplasmic reticulum in hepatic lipid homeostasis and stress signaling. Cell Metab. 15: 623–634. - PubMed
-
- Chiu H. C., Kovacs A., Blanton R. M., Han X., Courtois M., Weinheimer C. J., Yamada K. A., Brunet S., Xu H., Nerbonne J. M., et al. 2005. Transgenic expression of fatty acid transport protein 1 in the heart causes lipotoxic cardiomyopathy. Circ. Res. 96: 225–233. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
