

Type 1 Interferons Induce Changes in Core Metabolism that Are Critical for Immune Function
- PMID: 27332732
- PMCID: PMC5695232
- DOI: 10.1016/j.immuni.2016.06.006
Type 1 Interferons Induce Changes in Core Metabolism that Are Critical for Immune Function
Abstract
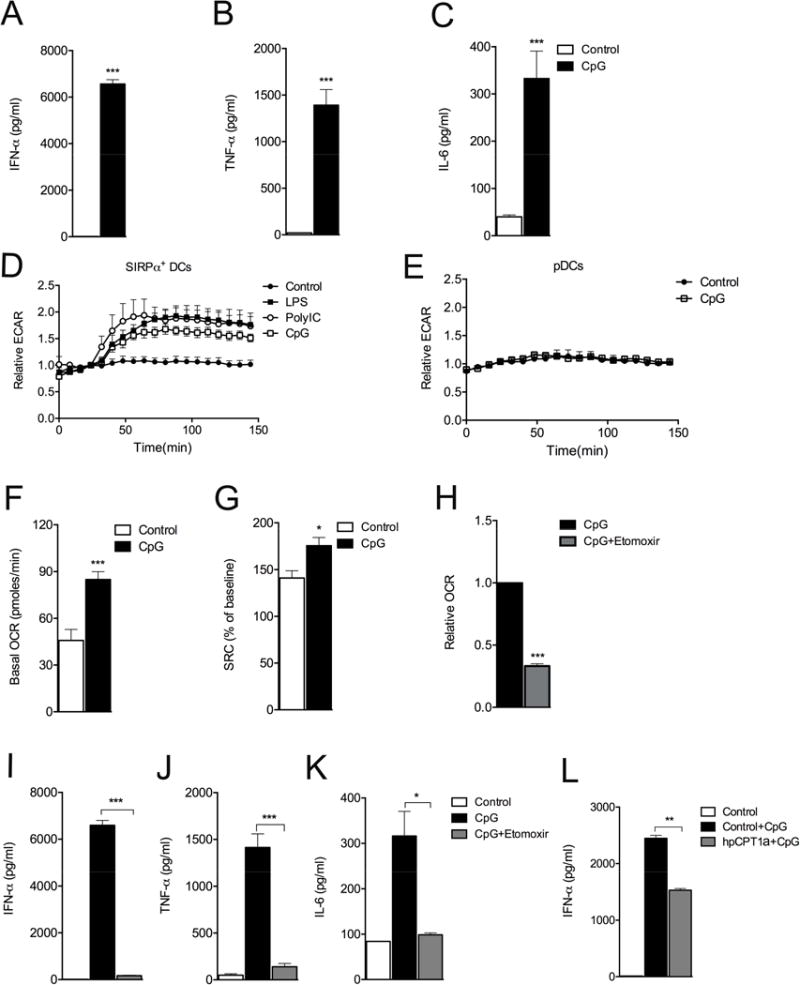
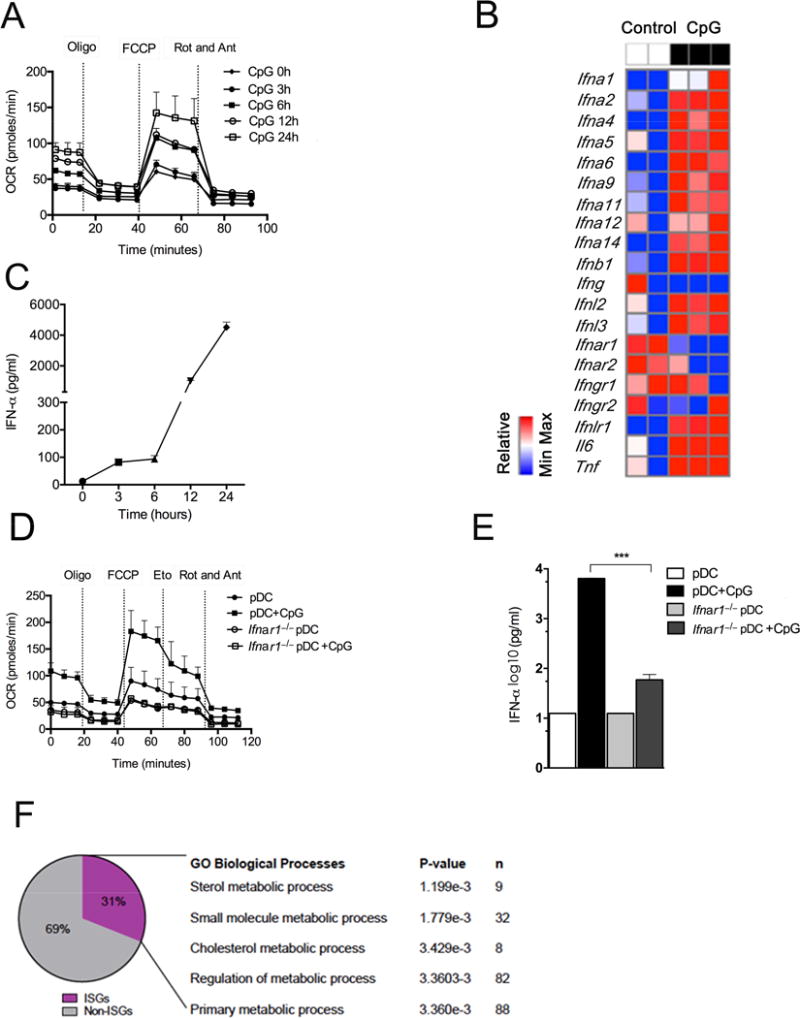
Greater understanding of the complex host responses induced by type 1 interferon (IFN) cytokines could allow new therapeutic approaches for diseases in which these cytokines are implicated. We found that in response to the Toll-like receptor-9 agonist CpGA, plasmacytoid dendritic cells (pDC) produced type 1 IFNs, which, through an autocrine type 1 IFN receptor-dependent pathway, induced changes in cellular metabolism characterized by increased fatty acid oxidation (FAO) and oxidative phosphorylation (OXPHOS). Direct inhibition of FAO and of pathways that support this process, such as fatty acid synthesis, prevented full pDC activation. Type 1 IFNs also induced increased FAO and OXPHOS in non-hematopoietic cells and were found to be responsible for increased FAO and OXPHOS in virus-infected cells. Increased FAO and OXPHOS in response to type 1 IFNs was regulated by PPARα. Our findings reveal FAO, OXPHOS and PPARα as potential targets to therapeutically modulate downstream effects of type 1 IFNs.
Copyright © 2016 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures





Comment in
-
pDCs Take a Deep Breath to Fight Viruses.Immunity. 2016 Jun 21;44(6):1246-8. doi: 10.1016/j.immuni.2016.06.005. Immunity. 2016. PMID: 27332726
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
