

Multiple Calmodulin-Binding Sites Positively and Negatively Regulate Arabidopsis CYCLIC NUCLEOTIDE-GATED CHANNEL12
- PMID: 27335451
- PMCID: PMC4981125
- DOI: 10.1105/tpc.15.00870
Multiple Calmodulin-Binding Sites Positively and Negatively Regulate Arabidopsis CYCLIC NUCLEOTIDE-GATED CHANNEL12
Abstract
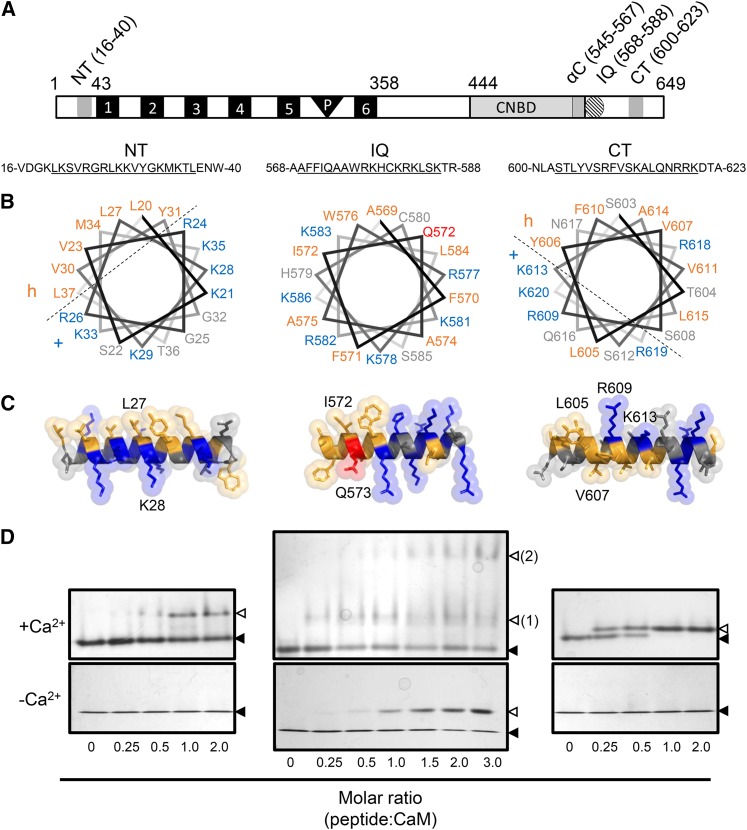
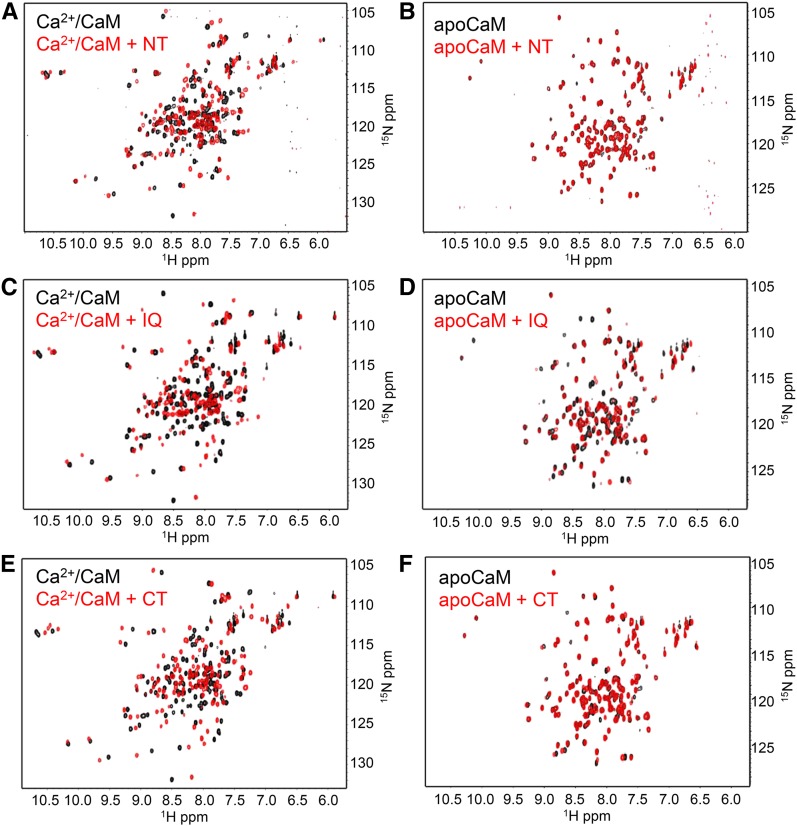
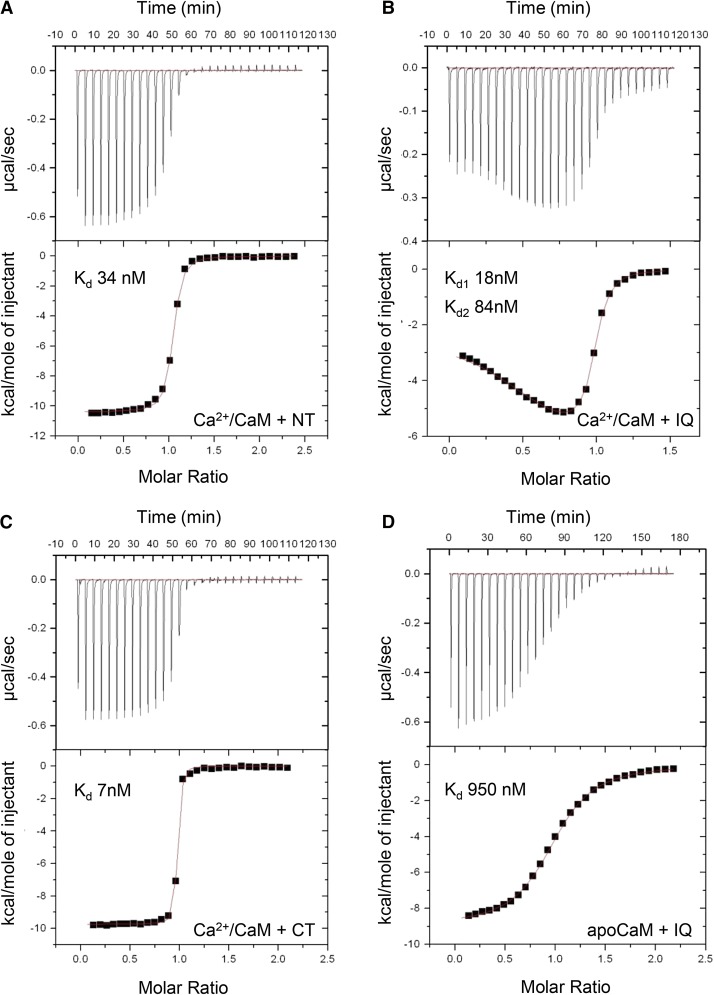
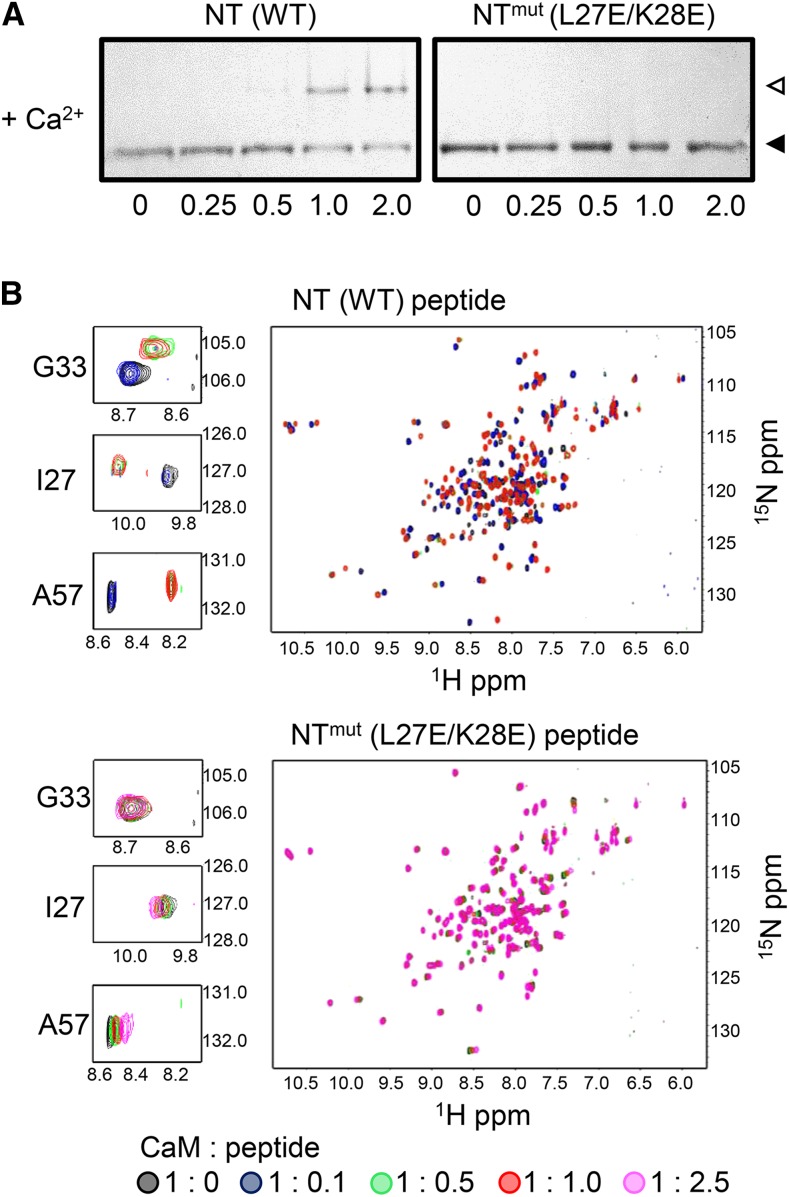
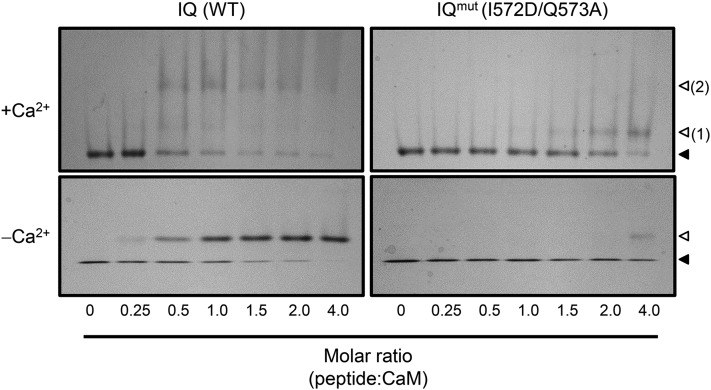
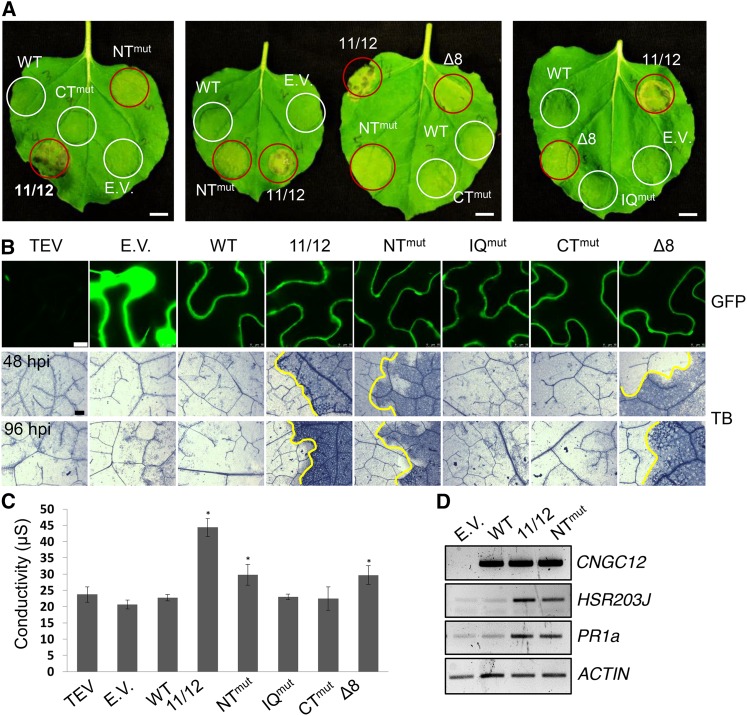
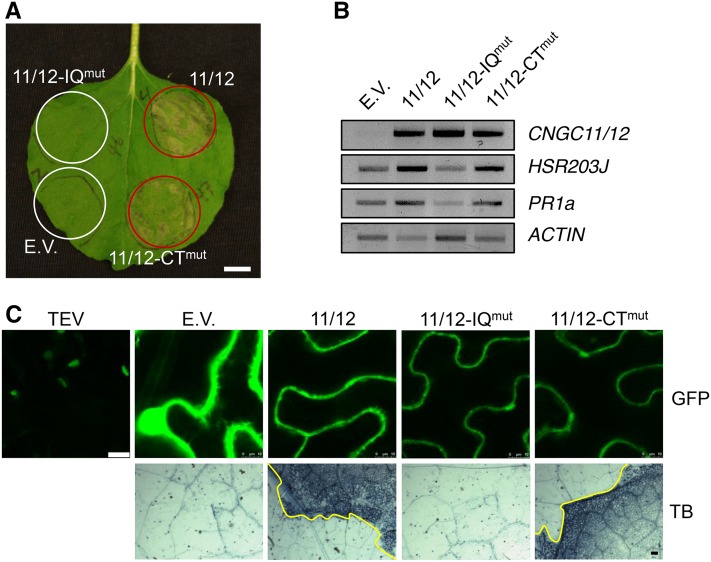
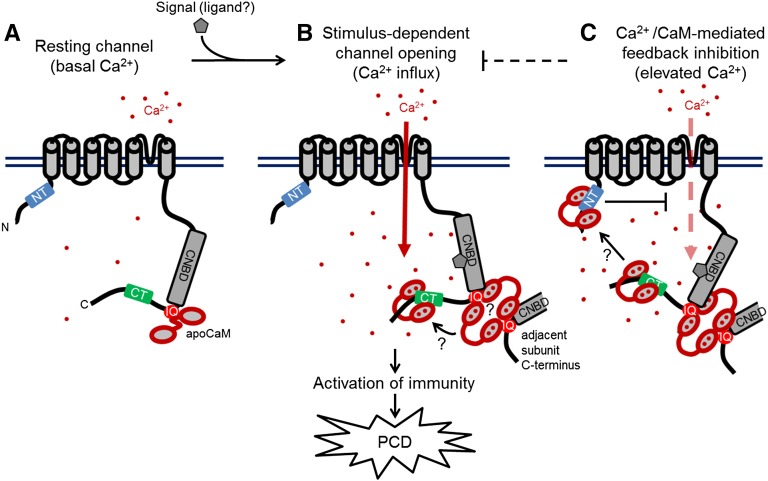
Ca(2+) signaling is critical to plant immunity; however, the channels involved are poorly characterized. Cyclic nucleotide-gated channels (CNGCs) are nonspecific, Ca(2+)-permeable cation channels. Plant CNGCs are hypothesized to be negatively regulated by the Ca(2+) sensor calmodulin (CaM), and previous work has focused on a C-terminal CaM-binding domain (CaMBD) overlapping with the cyclic nucleotide binding domain of plant CNGCs. However, we show that the Arabidopsis thaliana isoform CNGC12 possesses multiple CaMBDs at cytosolic N and C termini, which is reminiscent of animal CNGCs and unlike any plant channel studied to date. Biophysical characterizations of these sites suggest that apoCaM interacts with a conserved isoleucine-glutamine (IQ) motif in the C terminus of the channel, while Ca(2+)/CaM binds additional N- and C-terminal motifs with different affinities. Expression of CNGC12 with a nonfunctional N-terminal CaMBD constitutively induced programmed cell death, providing in planta evidence of allosteric CNGC regulation by CaM. Furthermore, we determined that CaM binding to the IQ motif was required for channel function, indicating that CaM can both positively and negatively regulate CNGC12. These data indicate a complex mode of plant CNGC regulation by CaM, in contrast to the previously proposed competitive ligand model, and suggest exciting parallels between plant and animal channels.
© 2016 American Society of Plant Biologists. All rights reserved.
Figures
References
-
- Arazi T., Kaplan B., Fromm H. (2000). A high-affinity calmodulin-binding site in a tobacco plasma-membrane channel protein coincides with a characteristic element of cyclic nucleotide-binding domains. Plant Mol. Biol. 42: 591–601. - PubMed
-
- Arazi T., Sunkar R., Kaplan B., Fromm H. (1999). A tobacco plasma membrane calmodulin-binding transporter confers Ni2+ tolerance and Pb2+ hypersensitivity in transgenic plants. Plant J. 20: 171–182. - PubMed
-
- Baxter J., Moeder W., Urquhart W., Shahinas D., Chin K., Christendat D., Kang H.G., Angelova M., Kato N., Yoshioka K. (2008). Identification of a functionally essential amino acid for Arabidopsis cyclic nucleotide gated ion channels using the chimeric AtCNGC11/12 gene. Plant J. 56: 457–469. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
