

Set1/COMPASS and Mediator are repurposed to promote epigenetic transcriptional memory
- PMID: 27336723
- PMCID: PMC4951200
- DOI: 10.7554/eLife.16691
Set1/COMPASS and Mediator are repurposed to promote epigenetic transcriptional memory
Abstract
In yeast and humans, previous experiences can lead to epigenetic transcriptional memory: repressed genes that exhibit mitotically heritable changes in chromatin structure and promoter recruitment of poised RNA polymerase II preinitiation complex (RNAPII PIC), which enhances future reactivation. Here, we show that INO1 memory in yeast is initiated by binding of the Sfl1 transcription factor to the cis-acting Memory Recruitment Sequence, targeting INO1 to the nuclear periphery. Memory requires a remodeled form of the Set1/COMPASS methyltransferase lacking Spp1, which dimethylates histone H3 lysine 4 (H3K4me2). H3K4me2 recruits the SET3C complex, which plays an essential role in maintaining this mark. Finally, while active INO1 is associated with Cdk8(-) Mediator, during memory, Cdk8(+) Mediator recruits poised RNAPII PIC lacking the Kin28 CTD kinase. Aspects of this mechanism are generalizable to yeast and conserved in human cells. Thus, COMPASS and Mediator are repurposed to promote epigenetic transcriptional poising by a highly conserved mechanism.
Keywords: Mediator; S. cerevisiae; cell biology; chromatin; chromosomes; epigenetics; genes; histone; human; nuclear architecture; transcription.
Conflict of interest statement
AS: Reviewing editor,
The other authors declare that no competing interests exist.
Figures
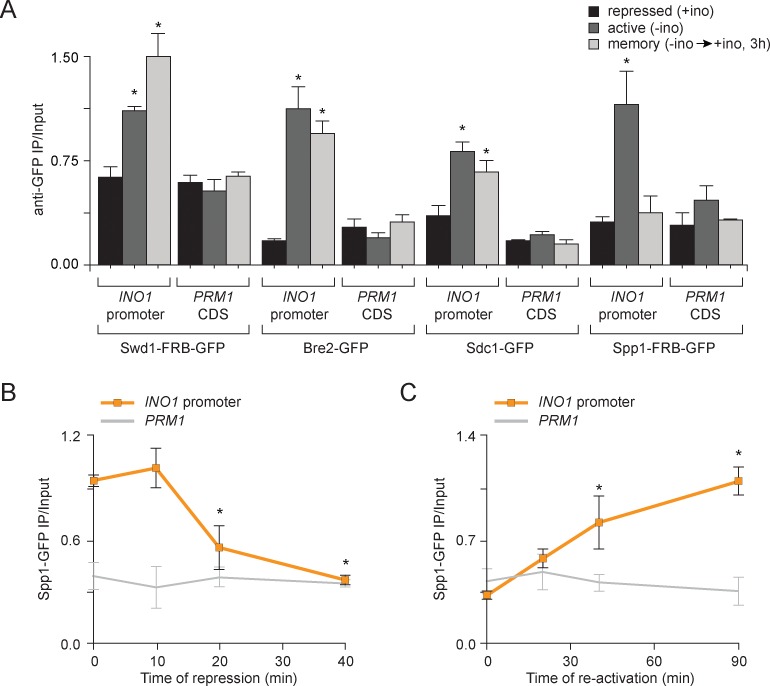
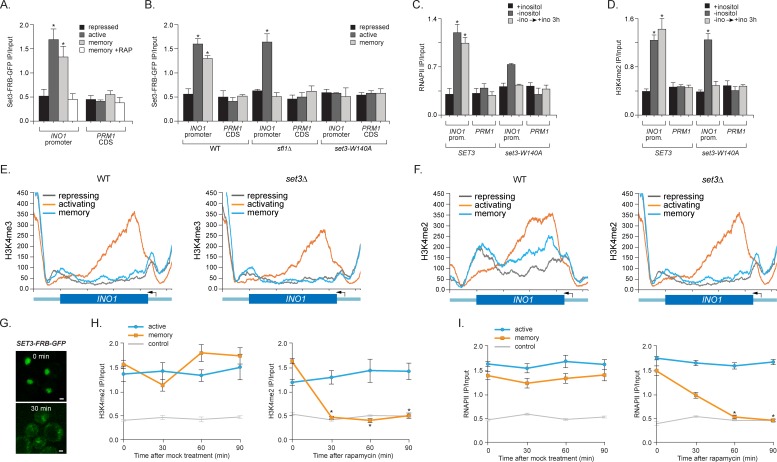
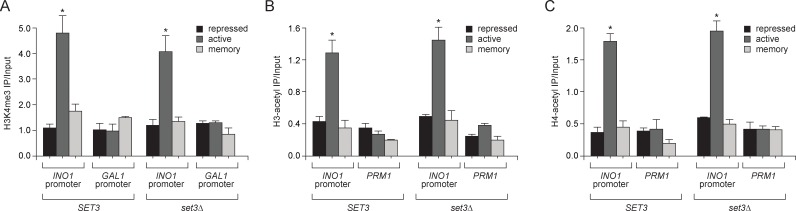
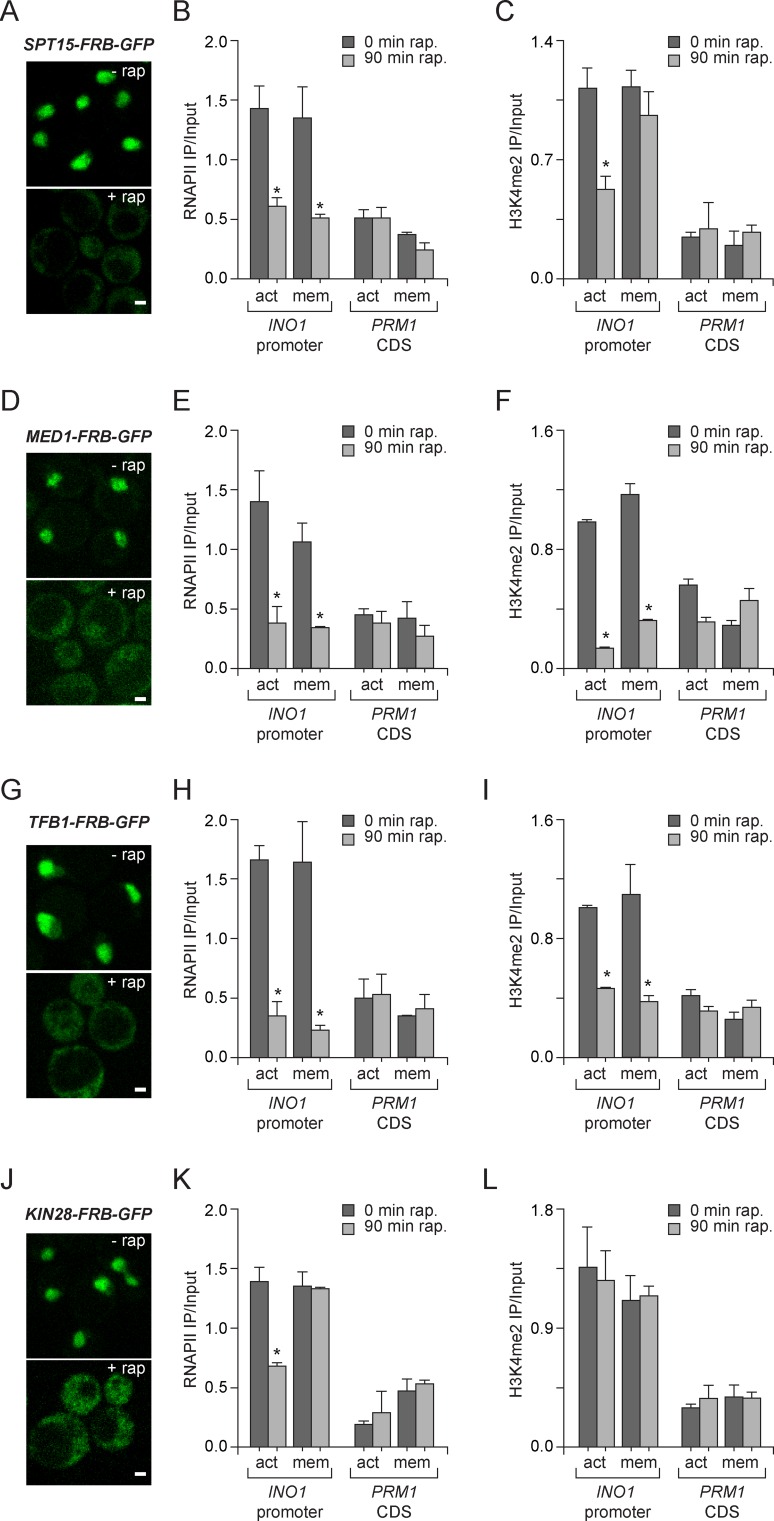
Similar articles
-
Mitotically heritable, RNA polymerase II-independent H3K4 dimethylation stimulates INO1 transcriptional memory.Elife. 2022 May 17;11:e77646. doi: 10.7554/eLife.77646. Elife. 2022. PMID: 35579426 Free PMC article.
-
The Saccharomyces cerevisiae Cdk8 Mediator Represses AQY1 Transcription by Inhibiting Set1p-Dependent Histone Methylation.G3 (Bethesda). 2017 Mar 10;7(3):1001-1010. doi: 10.1534/g3.117.039586. G3 (Bethesda). 2017. PMID: 28143948 Free PMC article.
-
Epigenetic Transcriptional Memory of GAL Genes Depends on Growth in Glucose and the Tup1 Transcription Factor in Saccharomyces cerevisiae.Genetics. 2017 Aug;206(4):1895-1907. doi: 10.1534/genetics.117.201632. Epub 2017 Jun 12. Genetics. 2017. PMID: 28607146 Free PMC article.
-
Establishment and inheritance of epigenetic transcriptional memory.Front Mol Biosci. 2022 Sep 2;9:977653. doi: 10.3389/fmolb.2022.977653. eCollection 2022. Front Mol Biosci. 2022. PMID: 36120540 Free PMC article. Review.
-
Nuclear pore proteins regulate chromatin structure and transcriptional memory by a conserved mechanism.Nucleus. 2013 Sep-Oct;4(5):357-60. doi: 10.4161/nucl.26209. Epub 2013 Aug 19. Nucleus. 2013. PMID: 23962805 Free PMC article. Review.
Cited by
-
Chromatin targeting of nuclear pore proteins induces chromatin decondensation.J Cell Biol. 2019 Sep 2;218(9):2945-2961. doi: 10.1083/jcb.201807139. Epub 2019 Jul 31. J Cell Biol. 2019. PMID: 31366666 Free PMC article.
-
Glucocorticoid signaling induces transcriptional memory and universally reversible chromatin changes.Life Sci Alliance. 2021 Aug 26;4(10):e202101080. doi: 10.26508/lsa.202101080. Print 2021 Oct. Life Sci Alliance. 2021. PMID: 34446533 Free PMC article.
-
Nuclear Control of Vascular Smooth Muscle Cell Plasticity during Vascular Remodeling.Am J Pathol. 2024 Apr;194(4):525-538. doi: 10.1016/j.ajpath.2023.09.013. Epub 2023 Oct 10. Am J Pathol. 2024. PMID: 37820925 Free PMC article. Review.
-
Rebelled epigenome: histone H3S10 phosphorylation and H3S10 kinases in cancer biology and therapy.Clin Epigenetics. 2020 Oct 14;12(1):147. doi: 10.1186/s13148-020-00941-2. Clin Epigenetics. 2020. PMID: 33054831 Free PMC article. Review.
-
Random sub-diffusion and capture of genes by the nuclear pore reduces dynamics and coordinates inter-chromosomal movement.Elife. 2021 May 18;10:e66238. doi: 10.7554/eLife.66238. Elife. 2021. PMID: 34002694 Free PMC article.
References
-
- Bancerek J, Poss ZC, Steinparzer I, Sedlyarov V, Pfaffenwimmer T, Mikulic I, Dölken L, Strobl B, Müller M, Taatjes DJ, Kovarik P. CDK8 kinase phosphorylates transcription factor STAT1 to selectively regulate the interferon response. Immunity. 2013;38:250–262. doi: 10.1016/j.immuni.2012.10.017. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
